Cell重磅综述:衰老的十二大特征
2023-02-16 MedSci原创 MedSci原创 发表于上海

衰老是一个永恒的话题,也是一个复杂的过程。随着年龄的增长,细胞的生长和分化能力以及生理功能都会逐步下降,导致个体的健康状况恶化,死亡风险增加。衰老还是许多常见疾病(如癌症、糖尿病、心脏病、神经退化等)
衰老是一个永恒的话题,也是一个复杂的过程。随着年龄的增长,细胞的生长和分化能力以及生理功能都会逐步下降,导致个体的健康状况恶化,死亡风险增加。衰老还是许多常见疾病(如癌症、糖尿病、心脏病、神经退化等)的根源。幸运的是,科学家们在衰老领域取得了令人惊叹的进展,特别是发现了一些可以用来判断和评估衰老程度的特征和标志物。2023年1月,在Cell杂志上发表了一篇重要的综述文章,题为“Hallmarks of aging: An expanding universe”,由西班牙奥维耶多大学的Carlos López-Otín和法国古斯塔夫鲁西研究所的Guido Kroemer等人撰写。这篇文章在2013年他们提出的九大衰老特征基础上,又增加了三个新发现的特征,形成了十二大衰老特征模型。这个模型为我们深入理解衰老机制以及开发有效延缓衰老方法提供了有力的指导。
事实上,这两位作者在2020年也在cell发表了重磅综述:重新定义健康,引发全球巨大的反响!同样这一篇重新定义衰老,也是极其重磅的!

作者认为衰老有以下十二个特征:基因组稳定性丧失、端粒损耗、表观遗传改变、蛋白稳态丧失、巨自噬障碍、营养感应失调、线粒体功能障碍、细胞衰老、干细胞耗竭、细胞间通讯改变、慢性炎症和生态失调。这些特征相互联系,也与最近提出的健康特征有关,包括空间分区的组织特征、内环境稳态、压力充分响应。
衰老研究主要探讨的是有机体在成年期的功能衰退。自2013年第一篇衰老特征综述在Cell上发表以来,已经发表了近三十万篇涉及衰老特征的文章,这与上个世纪的数量相当。因此,现在是整合十年间主要发现的新版衰老特征综述较为成熟的时机。
衰老特征是相互关联和影响的,而不是孤立存在的。所以,把它们分成不同的类别有些主观,但作者提出了三个通用的标准来判断一个特征是否属于衰老特征:(1)它会随着年龄发生变化;(2)它在实验中被增强会加速衰老;(3)它被干预治疗会延缓或逆转衰老。这篇文章没有详细描述与年龄相关的外在表现,而是重点介绍了分子、细胞和系统层面的过程,以及它们是如何导致衰老的。此外,在动物和人类中,要评估生物衰老情况,就需要对影响衰老个体形态和功能下降的因素进行客观量化。实际上,生物年龄和实际年龄之间的差异可以反映出促进或延缓衰老措施的效果,也可以评估某一特征在衰老过程中所起的作用。为了做到这一点,需要使用标准化的生理测量方法(比如测量基础和最大能量消耗)、功能测试方法(比如测试感觉、运动和认知能力)以及更先进的“组学”技术(比如基因组学、表观基因组学、转录组学、蛋白质组学和代谢组学),这些技术通常应用于单细胞水平,以便评估健康状况下降的时空模式和延缓衰老策略的效果。
2013年时,作者总结了九个分子、细胞和系统层面上的衰老特征:基因组稳定性丧失、端粒耗损、表观遗传改变、蛋白稳态丧失、营养感应失调、线粒体功能障碍、细胞衰老、干细胞耗竭和细胞间通讯改变。最新十年来的研究证实并强调了这些特征都是重要且有效的。它们已经经受住了无数衰老研究人员对其进行验证与检验,并且现在需要更新这些特征来反映最新十年来科学界取得成就。例如,在2013年时大多数延缓衰老措施只在非哺乳动物模型中得到证明,包括酵母菌、线虫和果蝇等。但幸运地是,在小鼠甚至非人灵长类动物中进行实验已经证明了这些措施也适用于哺乳动物。值得注意地是,在统计数据上显示如果人类与某一特征相关联并存在因果关系,则他们具有相似基因组特性并同时患有相关疾病概率更高,并且作者已经在临床上验证了他们提出推断。
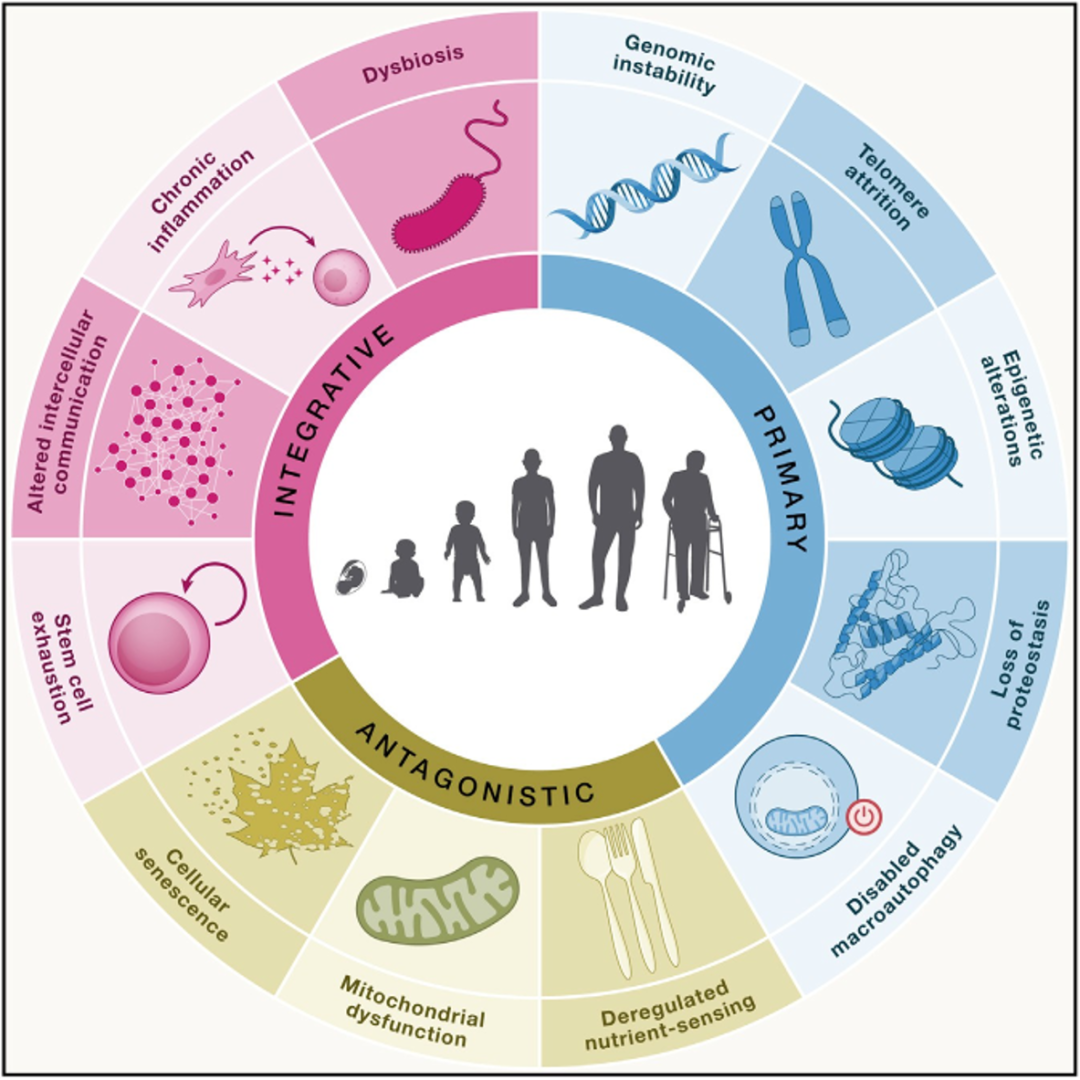
图1 衰老特征。该图汇集了本文提出的12个衰老特征: 基因组稳定性丧失、端粒耗损、表观遗传改变、蛋白稳态丧失、自噬障碍、营养感应失调、线粒体功能障碍、细胞衰老、干细胞耗竭、细胞间通讯改变、慢性炎症和微生态失调。这些特征可分为三类:原发性、拮抗性和综合性。
除了对以往的特征进行必要的更新之外,在这篇综述中作者还增添了一些内容,包括以下三个额外的衰老特征:自噬障碍、慢性炎症和微生态失调。巨自噬障碍最初被视为蛋白稳态失衡的特殊情况。然而,自噬不仅会影响蛋白质,还会靶向整个细胞器和非蛋白质大分子,因此有理由将其作为一个单独的特征进行讨论。此外,作者认为2013年列出的最后一个标志(细胞间通讯改变)太过宽泛,需要单独讨论慢性炎症和年龄相关的生态失调(图1)。
衰老特征的相互联系意味着一种特征的实验性加重或减弱通常也会影响其他特征。这表明了一个事实,即衰老是一个复杂的过程,必须作为一个整体来看待。因此,应将每一个特征视为未来探索衰老过程以及开发新延缓衰老药物的切入点。
一、基因组稳定性丧失
基因组是生物体的遗传蓝图,但它经常受到各种因素的破坏,比如化学物质、辐射、生物病原体、DNA复制错误、染色体分离错误、氧化应激和水解反应等。这些因素会导致基因组出现各种损伤,比如点突变、缺失、易位、端粒缩短、单链和双链断裂、染色体重排、核结构缺陷以及病毒或转座子的插入等。这些损伤会影响基因的功能和表达,从而引起生理和病理上的衰老。为了保护基因组免受损伤,生物体进化出了一套复杂的DNA修复和维护系统,可以修复细胞核DNA和线粒体DNA(mtDNA)的损伤,并保持染色体的结构和稳定性。然而,随着年龄增长,这些DNA修复系统会变得低效,导致基因组损伤和异常DNA在细胞内积累(图2A)。
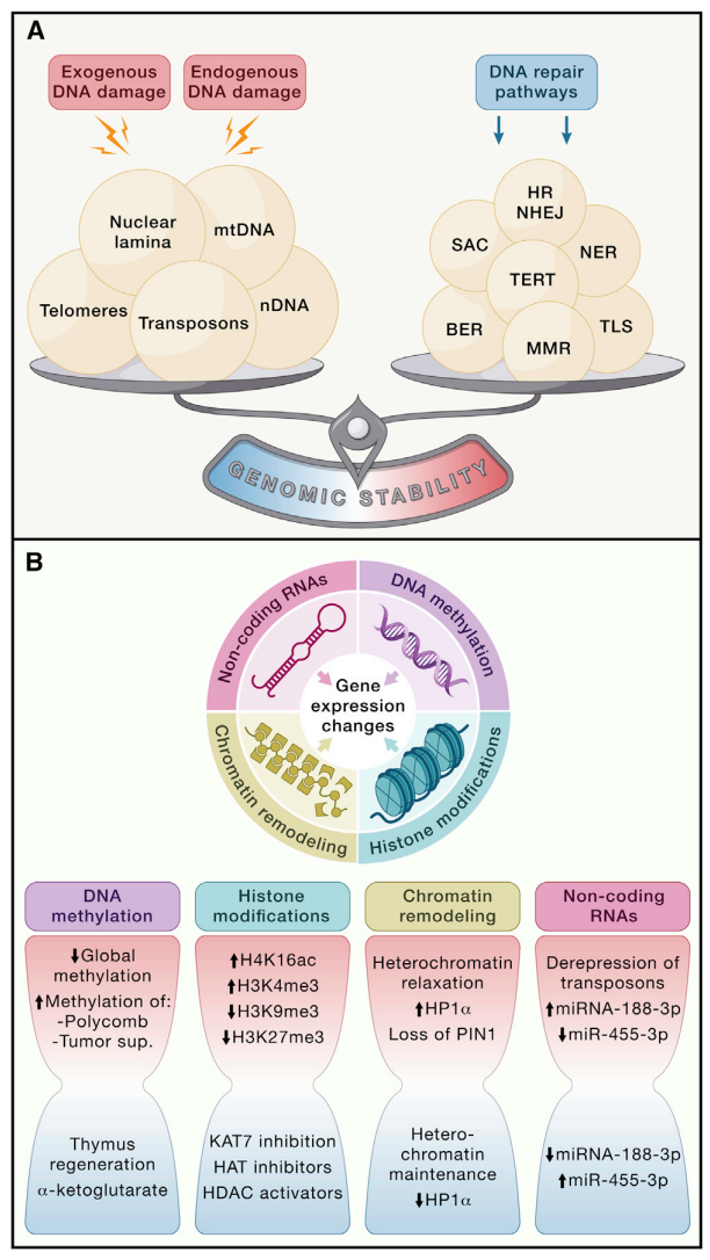
图2 基因组稳定性丧失、端粒损耗和表观遗传改变导致的细胞完整性丧失。(A)内源性或外源性因素会导致各种DNA损伤,从而导致生理性和病理性衰老。此类损伤可以通过多种机制进行修复,但这些机制往往会随着年龄增长而降低效率。DNA过度损伤、DNA修复不足、核结构改变和端粒磨损会促进衰老过程。BER,碱基切除修复;HR,同源重组;NER,核苷酸切除修复;NHEJ,非同源末端连接;MMR,错配修复;SAC,纺锤体组装检查点;TERT,端粒酶逆转录酶;TLS,跨损伤DNA复制。(B)DNA或组蛋白的乙酰化和甲基化变化,以及染色质相关蛋白或非编码RNA(ncRNA)的水平或活性的变化,会诱发导致衰老的表观遗传变化。沙漏的红色部分表示衰老相关的变化,蓝色部分表示避免它们的策略。
拓展:
核DNA
基因组损伤是衰老的重要原因之一。生物体的细胞核DNA会受到外源和内源因素的影响,产生各种形式的损伤,如突变、缺失、易位、断裂、重排等。这些损伤会干扰基因的功能和表达,导致细胞功能失调和组织稳态破坏。特别是当干细胞受到DNA损伤时,会影响其自我更新和分化能力,从而加速衰老和增加疾病风险。特别是,当DNA损伤影响干细胞,即阻碍它们的自我更新或导致它们耗竭时,又会促进衰老并增加对衰老相关疾病的易感性。组织学上正常的人体组织中的突变负荷是显著的。例如,年轻个体的正常食管上皮细胞已经显示出数百个突变,到中年时每个细胞可能携带2,000多个突变。整个生命过程中DNA突变的积累可能是可以容忍的,因为由外源性和内源性因素引起的所有基因组损伤的完全修复的能量成本过高。因此,细胞更倾向于生存而不是保持基因组完整。这些数据还表明,人体组织中的突变负荷随着年龄增长而增加,但并不一定导致衰老或癌症,因为它们还需要一个适合的微环境才能发挥作用。
不同物种之间的比较表明,体细胞突变率与寿命呈负相关。生物体进化出了一套复杂的DNA修复系统来应对基因组损伤,但随着年龄增长,这些系统会变得低效。有证据表明,DNA修复机制的缺陷或改变会加速或延缓衰老,并与一些人类早衰综合征有关。因此,提高DNA修复效率或减少基因组损伤可能是延缓衰老和预防疾病的有效策略。迄今为止,尽管尚无明确证据表明正常的突变固定率是导致衰老的原因,但大量研究表明DNA修复缺陷有可能导致衰老。DNA修复机制的改变加速了小鼠的衰老,并成为几种人类早衰综合征的基础。相反,过表达有丝分裂检查点激酶BubR1的转基因小鼠表现出健康寿命的延长(表1)。此外,对人类和其他长寿物种的研究表明,增强的DNA修复机制会随着寿命的延长而共同进化。Sirtuin-6(SIRT6)可能在不同物种的修复效率中发挥重要作用。小鼠中SIRT6的过表达降低了基因组不稳定性,改善了双链断裂修复,并延长了寿命(表1),尽管其他证据,例如改善葡萄糖代谢和恢复能量稳态,也可解释SIRT6的延长寿命效应。值得注意的是,最近的研究表明,通过小分子激活8-氧鸟嘌呤DNA糖基化酶1促进了氧化性DNA损伤的修复,并且可能在衰老和与过度氧化损伤相关的其他过程中具有治疗应用。这些发现表明,旨在减少核DNA突变负荷或者增强/改变其修复机制的干预措施可能会延缓衰老和衰老相关疾病的发作,但在这方面仍然缺乏进一步的因果证据。
拓展:Cell | 刘光慧/曲静/张维绮揭示古病毒复活驱动程序性衰老
线粒体DNA
影响mtDNA不稳定性可能会导致衰老和衰老相关的病理学。mtDNA由于其高复制率、修复机制效率有限、线粒体内氧化微环境,以及缺乏保护性组蛋白,尤其会受到衰老相关突变和缺失的影响。在衰老过程中,人体组织的mtDNA突变增加,但尚不清楚这种增加是否在功能水平影响衰老过程。由于“异质性”,即同一细胞内突变基因组和野生型基因组共存,mtDNA突变在驱动衰老中的因果关系很难评估。然而,对衰老细胞的深度测序表明,它们的mtDNA突变负荷可能会通过克隆扩增事件显著增加。此外,在灵长类动物卵母细胞和组织中、神经退行性疾病患者的淋巴母细胞中,也观察到线粒体突变随年龄的增长而加速增加。值得注意的是,超灵敏测序表明,衰老细胞中的大多数mtDNA突变是由mtDNA聚合酶γ引起的复制错误导致的,而不是由氧化应激导致的。
mtDNA突变可以引发疾病,并且部分表型和衰老相似,这些证据初步表明mtDNA突变可能直接参与衰老和衰老相关疾病。进一步的证据来自对缺乏DNA聚合酶γ的小鼠的研究,这些小鼠表现出加速衰老和mtDNA缺失而不是点突变相关的寿命缩短(表1)。总体而言,这些数据表明,避免、减弱或纠正mtDNA突变可能有助于延长健康寿命和预期寿命。然而,与核DNA突变的情况一样,证明促进mtDNA修复机制中功能可以延缓衰老的实验证据仍然在很大程度上缺失。
拓展:PNAS:顾正龙团队揭示衰老个体中线粒体DNA突变广泛存在且高度有害
cells:研究发现补铁可通过增强线粒体功能延缓衰老并延长寿命
细胞核结构
核纤层构成了锚定染色质和蛋白质复合体的支架,其缺陷会导致基因组不稳定。例如,Hutchison-Gilford早衰症(HGPS)和Nestor-Guillermo早衰症(NGPS)等加速衰老综合症就是由编码核纤层蛋白组分的LMNA基因和BANF1基因突变引起的。除此之外,核纤层的改变以及Lamin A前体异构体progerin的产生也是正常人类衰老过程中的显著特征,并且在细胞衰老的过程中也可以看到Lamin B1水平的降低。动物和细胞模型已经促进了对于衰老和早衰引起的核纤层异常引发的应答机制和应激途径的识鉴定,其中包括肿瘤抑制因子p53(TP53)的激活,生长轴的失调以及成体干细胞的损耗等。
在HGPS小鼠模型中,通过全身注射反义寡核苷酸,法尼基转移酶抑制剂,他汀类药物和氨基二膦酸盐的混合剂,修复生长轴或阻断NF-κB通路,可以降低Lamin A前体或progerin的水平,并可以延缓早衰症状的产生同时延长小鼠的生命周期。这证明了核纤层异常与早衰之间的因果关系,且这些干预措施中的一部分已经被批准用于早衰症患者的临床治疗中了。除此之外,最近开发的基因编辑技术已经被用于修正HGPS患者细胞和该疾病动物模型中的LMNA基因突变。在未来,这些方法有望投入到早衰症的临床治疗中,但到目前为止,仍没有任何证据表明减少progerin可以延缓正常衰老。
二、端粒耗损
染色体末端(端粒)的DNA损伤会导致衰老以及衰老相关疾病。端粒损伤会导致DNA复制酶无法完成端粒区域的真核DNA复制,因此在几轮细胞分裂后,端粒大幅度缩短从而诱发基因组不稳定,并最终导致细胞凋亡或衰老。这些有害的影响可以通过端粒酶的逆转录活性来消除,端粒酶是一种活性核糖核蛋白,其可以延长端粒来保证它们具有足够的长度。然而,大多数哺乳动物体细胞并不表达端粒酶,这导致随着细胞寿命的增加,端粒序列会受到持续不断的累积性损伤。有研究表明,端粒损耗可以通过限制癌细胞的复制来减缓癌变过程。因此,与基因组不稳定性明显有利于肿瘤发生相反,端粒损耗可能具有拮抗恶性肿瘤的作用。出于这个原因,作者认为端粒损耗是一个独立于基因组不稳定性的衰老标志。
人类端粒酶缺乏与肺纤维化,再生障碍性贫血和先天性角化不良等疾病的过早发生有关,所有这些疾病都阻碍了受影响组织的再生能力。端粒缩短也在包括人类和小鼠在内的多个物种的衰老过程中被观察到。端粒损耗率受到年龄,遗传变异,生活方式和社会因素的影响,且取决于受影响细胞的增殖活力,并可以用于预测许多物种的寿命长度。端粒脱帽也可能是缺乏端粒蛋白复合体的结果,端粒蛋白复合体是一类具有阻断染色体末端DNA损伤反应并且调节端粒长度能力的蛋白。端粒蛋白复合体的几种功能丧失模型表明,即使端粒长度正常,组织的再生能力也会下降且表现出加速衰老。
转基因动物模型解释了端粒损耗、细胞衰老和机体衰老之间的因果关系。端粒缩短或延长的小鼠分别表现出寿命的缩短或延长。值得注意的是,当端粒酶被基因编辑重新激活后,端粒酶缺陷小鼠的早衰症状可以被逆转(表1)。除此之外,通过药物激活或者病毒激活端粒酶可以延缓小鼠的正常衰老,而端粒超长的小鼠则显示出寿命的延长和代谢健康的改善(表1)。类似地,在阿尔茨海默症的小鼠模型中,经过改造使得成体神经元可以维持生理水平端粒酶的小鼠表现出神经元的存活以及认知功能的维持(表1)。因此,衰老可以通过控制端粒酶的激活来调节。
Genes Devel:科学家发现控制细胞衰老的开关—端粒酶
端粒酶活化可以延缓衰老并治疗端粒疾病
在人类中,许多研究已经证明了端粒长度缩短与衰老相关疾病之间的因果关系。尤其是利用端粒缩短的小鼠模型已经证明端粒损耗是端粒综合征和主要的衰老相关疾病(如肺和肾纤维化)的原因。这些端粒动力学和机体衰老之间的联系使得研究人员可以设计新的干预措施来延缓衰老以及衰老相关疾病。例如,使用基因治疗方法激活端粒酶的策略已经在肺纤维化和再生障碍性贫血的小鼠模型中取得了治疗效果。
三、表观遗传学的改变
导致衰老的各种表观遗传变化包括DNA甲基化模式的改变,组蛋白翻译后修饰异常,异常染色质的重塑以及非编码RNA(ncRNAs)的功能失调等(图2B)。这些调节和可逆的变化影响了基因表达和其他细胞活动,并最终引起了癌症、神经退行性疾病、代谢综合征以及骨科疾病等一些与衰老相关的疾病的发展和恶化。大量的酶系统地参与了表观遗传模式的产生与维持,这些酶包括DNA甲基转移酶、组蛋白乙酰化酶、去乙酰化酶、甲基化酶,去甲基化酶以及涉及染色质重塑或ncRNA合成和成熟的蛋白复合体。
DNA甲基化
人类DNA甲基化修饰的模式随着时间的推移积累了多种多样的变化。早期研究描述了与衰老相关的整体低水平甲基化,但进一步的分析显示包括一些肿瘤抑制基因和Polycomb靶基因在内的特定位点会随着衰老过程甲基化水平逐渐升高。早衰综合症患者和小鼠的细胞也表现出DNA甲基化的变化,这一变化部分重现了正常衰老过程中发现的甲基化改变。但是,由于大多数这些与衰老相关的表观突变影响了内含子和基因间区域,导致这些表观突变产生的功能结果是不确定的。
基于特定部位DNA甲基化状态的表观遗传时钟已经被用于预测实际年龄和死亡风险,以及评估可能延长人类寿命的干预措施。针对胸腺再生的方案已经证明了这一点,实施该方案后,许多与衰老相关疾病风险指数得到改善。同时在1年的治疗后,患者平均表观遗传年龄比基线低1.5年左右。此外,人类发病率和死亡率的预测显示,表观遗传年龄比实际年龄低2年,并且这一状况一直持续到停止治疗6个月后。类似地,连续补充α-酮戊二酸7个月也使得表观遗传时钟拨回了8年。综上所述,DNA甲基化变化与衰老有关,但是尚没有确切证据表明其会导致衰老。目前仍需要进一步研究来证明DNA甲基化维持的缺陷是否会导致衰老,以及DNA甲基化维持的保真度提高能否延长寿命。鉴定负责调控衰老过程中DNA甲基化发生变化的分子驱动因素是必须的。
拓展:Neurology:血液DNA甲基化测定的衰老速度与年龄相关性认知障碍和痴呆的关系
组蛋白修饰
组蛋白的整体丢失以及组蛋白翻译后修饰的组织依赖性变化也与衰老密切相关。组蛋白表达的增加延长了果蝇的寿命,而组蛋白H4K16ac或者H3K4me3水平的增加以及H3K9me3或H3K27me3水平的降低在老年人和早衰症患者的成纤维细胞中被发现。这些组蛋白修饰会导致转录本改变,细胞稳态丢失以及衰老相关代谢水平的下降。值得注意的是,端粒上异染色质标记的丢失已被证明会导致端粒的延长。
组蛋白去甲基化酶通过靶向关键长寿路径的成分(如胰岛素/胰岛素样生长因子-1(IGF-1)信号通路)调节寿命。其他组蛋白修饰酶,例如蛋白质去乙酰化酶SIRT家族成员和ADP-核糖转移酶也有助于健康衰老。尽管没有增加寿命,但过表达SIRT1提高了小鼠衰老过程中的基因组稳定性和代谢效率。过表达SIRT3可以逆转衰老造血干细胞(HSC)丧失的再生能力,并介导了利于长寿的饮食限制的有益影响。类似地,小鼠体内Sirt6敲除会导致加速衰老,而Sirt6过表达则可以延长寿命。其影响寿命的机制是由于Sirt6是一种多功能蛋白,具有将染色质动力学与代谢和DNA修复相互连接的能力。最后,Sirt7缺失会导致全基因组不稳定,代谢功能障碍和早衰。总之,这些发现与去乙酰化酶活性降低会导致染色质松弛,DNA损伤增加和基因组不稳定性增加的观点是一致的。相反地,人类干细胞中组蛋白乙酰转移酶KAT7失活降低了组蛋白H3K14ac,并延缓了细胞衰老的表型。此外,静脉注射编码Cas9/sg-Kat7的的慢病毒可以改善正常小鼠和早衰小鼠的肝细胞和肝脏衰老,并延长了两种小鼠的寿命。抑制组蛋白乙酰转移酶可以改善早衰症小鼠的表型并延长其寿命,而组蛋白去乙酰化酶激活剂可以部分地通过上调SIRT1活性来延长寿命。总之,这些发现表明,组蛋白修饰子应该被作为部分延缓衰老相关的治疗策略的一部分进一步深入研究,尽管尚不清楚这些干预措施是通过纯粹的表观遗传机制,通过影响DNA修复和基因组稳定性还是通过影响代谢或信号通路的转录变化来影响衰老和寿命。
染色质重塑
除了DNA和组蛋白修饰,一些染色体蛋白和染色质重塑因子,例如影响基因组稳定性和DNA修复的异染色质蛋白1α(HP1α)和Polycomb家族蛋白,可能具有调节衰老的作用。这些表观遗传因子的改变可能会导致包括全基因组异染色质丢失和重新分配在内的染色质结构改变,而这在衰老细胞中也很常见。
这些染色质改变在衰老过程中的因果关系已经在无脊椎动物中被大量研究,例如HP1α的功能丧失会减少寿命,而其过表达则会促进健康并延长寿命(表1)。虽然在哺乳动物中类似的研究仍然有限,但大多数研究已经表明异染色质松弛会导致衰老和衰老相关疾病,而异染色质的维持则会促进长寿。例如,在从果蝇到哺乳动物的不同物种中,对于保持异染色质结构所必须的PIN1-脯氨酸异构酶的缺失与早衰和神经退行性病变有关(表1)。然而,仍然缺少通过调节染色质重塑因子的功能来延长脊椎动物寿命的实验。
非编码RNAs
数量庞大且不断增长的ncRNAs群体,包括长链非编码RNA(long noncoding RNAs,IncRNAs,如端粒RNAs或TERRA),微RNA(microRNAs,miRNAs) ,和环状RNAs已成为可以影响衰老进程的表观遗传因子。ncRNAs通过靶向转录后的长寿网络组分或通过调节干细胞行为调节健康寿命和寿命。一种环状RNA调节胰岛素/IGF-1信号通路来影响果蝇寿命,但大多数研究都集中在miRNAs上,对于其他ncRNA可能在多大程度上源自转录噪声仍存在争议,其在人类生理和病理中的调节作用仅限于少数特定情况。
功能增益和功能缺失研究首先证实了几种miRNAs调节无脊椎动物寿命的能力。对小鼠的后续研究提供了miRNAs在衰老中的功能相关性的因果证据(表1)。例如,miRNA-188-3p随衰老在骨骼内皮中表达上调,并导致与衰老相关的血管问题。小鼠中miR-188的缺失可以延缓对骨有益的毛细血管的随衰老减少,而miR-188的内皮特异性过表达可以降低骨质量并抑制骨再生。相反,小鼠中miR-455-3p的缺失表现出对线粒体动力学、认知行为和寿命的有害效果,而它的过表达保留了这些功能并延长寿命。总的来说,这些发现表明miRNAs可能导致衰老和衰老相关病理,并可作为延缓衰老及衰老相关疾病的潜在治疗靶点。
逆转录转座子的去抑制
最近的研究揭示了逆转录转座子在包括人类在内的复杂后生动物衰老过程中的作用。这些逆转录转座元件是可移动的遗传单元,可以使用一种涉及RNA中间体的分子机制,从一个基因组位置移动到另一个。逆转录转座子由长散在核元件(LINEs)和短散在核元件(SINEs)组成,前者编码逆转录转座所需的蛋白质,后者是短的非编码RNAs,劫持LINE蛋白机器。逆转录转座子在衰老细胞中被重新激活,并通过遗传和表观遗传变化或识别逆转录转座子核酸为外源DNA后激活免疫通路的激活而产生有害影响。对LINE-1 RNA的表观遗传去抑制抑制了表观遗传阅读器Suv39H1,2导致H3K9me3和异染色质的整体减少,而LINE-1 RNA的逆转录产生双链cDNA,激活cGAS/STING/干扰素通路。
核苷类逆转录酶抑制剂(NRTIs)抑制或减弱逆转录转座,可延长Sirt6缺失小鼠的寿命,改善健康寿命,改善骨骼和肌肉表型(表1)。同样,用NRTIs治疗老年野生型小鼠可降低DNA损伤标记物的水平。此外,在体内用反义寡核苷酸靶向逆转录转座子可以增加早衰小鼠的寿命。值得注意的是,百岁老人中一种罕见的SIRT6变体是LINE1逆转录转座子的更强抑制因子,增强了基因组的稳定性,并且可以比野生型SIRT6更强有力地杀死癌细胞。综上所述,这些发现表明逆转录转座子促进了衰老过程,而对抗逆转录转座子活动的干预措施可能会改善健康长寿效果。针对逆转录转座子不同功能的药物在老年人群中的进一步临床研究可能会给出针对衰老和衰老相关疾病的新干预策略。
拓展:Science Translational Medicine:靶向LINE-1 RNA可改善早衰综合征的衰老表型
基因表达变化
以上这些表观遗传因子的作用机制主要集中在基因表达水平的调控上。衰老导致转录噪声的增加和许多mRNAs的异常生成和成熟。对人类和其他物种年轻和年老组织基于基因芯片的比较分析已经确定了衰老相关的转录特征,这些转录特征是由衰老过程中发生的表观遗传变化引起的。环境暴露也可以通过调控DNA甲基化和组蛋白修饰引起基因调控的改变,并促进与衰老相关的表观遗传变化,包括表观遗传时钟的加速。
在整个小鼠生命周期中不同年龄的多种细胞类型和器官的单细胞转录组学和血浆蛋白质组学揭示了衰老过程中显著变化的基因。这些变化特异地影响某些生物学过程,如炎症、蛋白质折叠、细胞外基质(ECM)调节和线粒体功能,这些过程在衰老过程中逐渐失调。不同组织在衰老过程中被观察到的共同表达模式可能有助于未来指导开发旨在改善健康寿命和寿命的干预措施(表1)。同样,在衰老过程中观察到的转录和转录后效率和保真度的下降,及其对蛋白质组的负面影响也可能为延长寿命策略的开发带来新的机遇。
四、蛋白质稳态丢失
衰老和一些衰老相关疾病,如肌萎缩性侧索硬化症(ALS)、阿尔茨海默症、帕金森病和白内障,与蛋白质稳态受损有关,导致错误折叠、氧化、糖化或泛素化蛋白质的积累,这些蛋白质通常形成聚集体,如细胞内包涵体或细胞外淀粉样斑块。
蛋白质稳态崩溃
由于错误翻译、错误折叠或不完整蛋白质的产生增多,细胞内蛋白质稳态可能被破坏(图3)。通过对核糖体蛋白RPS23的基因操纵来提高RNA到蛋白质翻译的准确性,可以延长酵母菌、秀丽隐杆线虫和黑腹果蝇的寿命,而一种导致翻译错误的RPS9突变,会致使小鼠过早衰老。另一种导致蛋白质稳态崩解的机制是蛋白质翻译延伸的减慢和氧化损伤的积累,这会分散用于蛋白质折叠的分子伴侣而蛋白质的正常折叠是维持细胞正常功能所必须的。此外,许多与年龄有关的神经退行性疾病,包括渐冻症和阿尔茨海默症,都可能是由蛋白质突变引起的,这些蛋白质突变使蛋白容易发生错误折叠和聚集,从而影响维持健康状态所需的蛋白质修复、清除和更新机制。
当保证质量控制的机制失效时,蛋白质稳态网络也会崩解,例如由于内质网(ER)中未折叠蛋白反应(UPR)功能下降,正确折叠的蛋白质的稳定性受损或用于降解蛋白质的蛋白酶体或溶酶体机制变得不足时(图3)。例如,在包括短寿命的弗氏假鳃鳉的大脑等衰老的器官中观察到蛋白酶体活性的降低。此外,一些单泛素化蛋白在苍蝇、小鼠、猴子和人类的衰老组织中积累,如组蛋白2A。
溶酶体对蛋白质的降解可以通过分子伴侣介导的自噬(CMA)以特定的方式实现,其中暴露类似KFERQ的五肽模体的蛋白质首先与热休克蛋白HSC70结合,然后与溶酶体相关的膜蛋白2A (LAMP2A)结合,这有助于将靶蛋白转位到溶酶体的腔内。小鼠肝脏LAMP2A的表达随着年龄的增长而下降,促进其表达可延缓肝脏衰老。蛋白质聚集物也可以包含在双膜囊泡,即自噬体中,随后与溶酶体融合而被巨自噬去除。由于自噬体可以包裹非蛋白质的细胞结构,这一过程将与蛋白质稳态分开在后面讨论。然而,刺激自噬构成了一种有效的清除细胞内蛋白质聚集物的策略。
拓展:Nature重磅发现:衰老的根源在核糖体?衰老加剧核糖体暂停,破坏蛋白质稳态
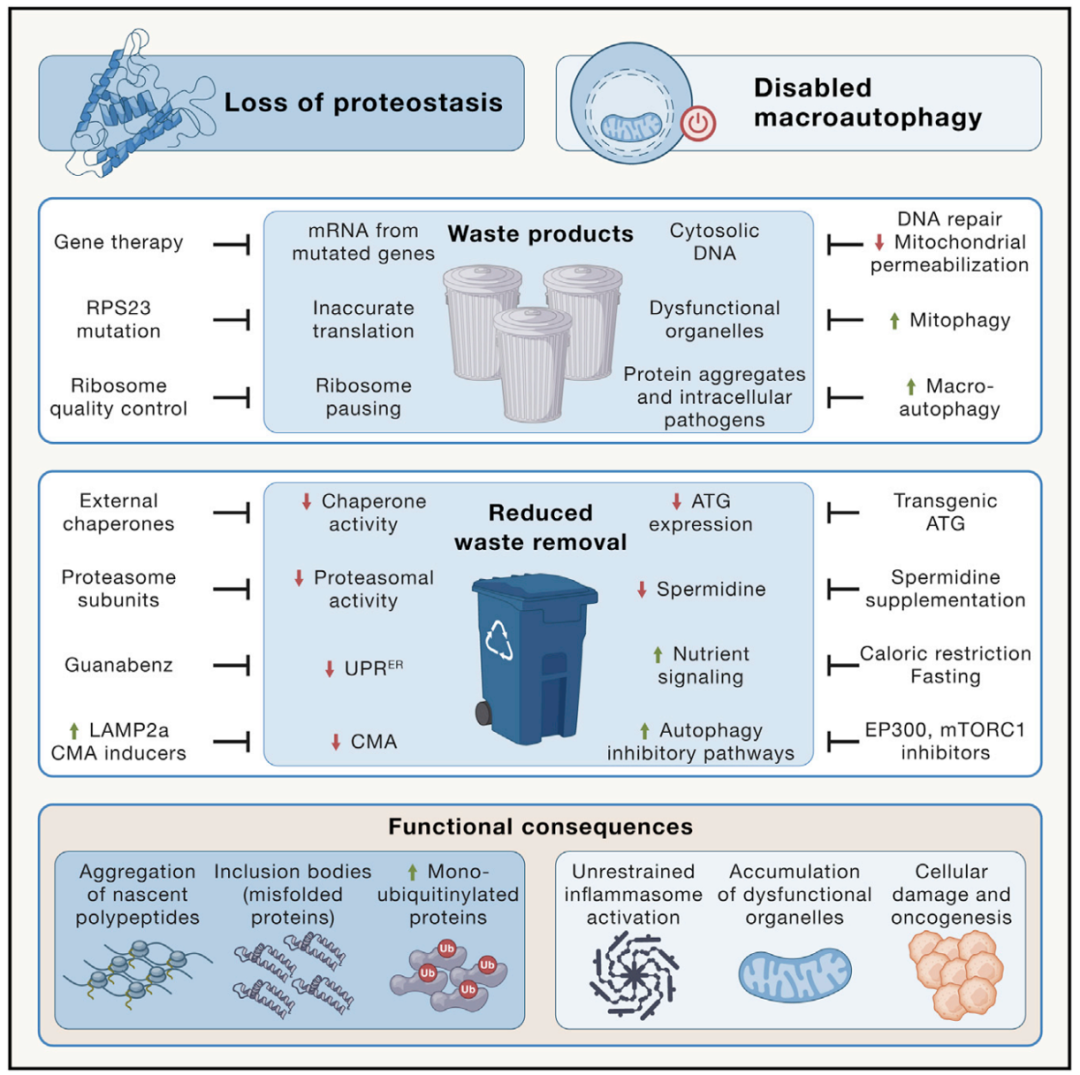
图3 蛋白质与细胞器的周转功能丢失。蛋白质稳态和巨自噬功能丧失的特征是偏离年轻平衡状态,其中废物的积累是由各种年龄相关的改变引起的,同时废物的清除通过各种机制受到损害。这些改变对功能的影响已列举。左图和右图举例说明了一些重建蛋白质稳态和自噬的策略。
蛋白质稳态,衰老与长寿
蛋白质稳态的剧烈扰动会加速衰老。例如,用糖基化终产物(AGEs)或脂褐素(共价交联蛋白质、糖和脂质的聚集物)喂养果蝇会导致AGE修饰和羰基化蛋白质的积累,从而缩短健康寿命和寿命,这在溶酶体蛋白酶组织蛋白酶D被敲低时进一步加剧。蛋白酶ZMPSTE24的缺失破坏了prelamin A的正常成熟,并在小鼠中引起早衰综合征,在ZMPSTE24功能缺失突变的人类中已观察到相似表型。在小鼠中,敲除神经元中的LAMP2A(对CMA至关重要)会深刻地影响蛋白质组,产生与阿尔茨海默症患者相似的变化。事实上,在小鼠中抑制CMA会加剧实验性阿尔茨海默症,而CMA激活剂则会缓解疾病表型。
通过实验改善蛋白稳态可延缓衰老进程(表1)。小鼠经鼻内应用重组人HSP70蛋白可增强蛋白酶体活性,降低脑脂褐素水平,增强认知功能,延长寿命。同样地,给老年小鼠服用化学分子伴侣4-苯基丁酸盐可以减少大脑中的内质网压力,提高认知能力。在线虫和果蝇中,过表达蛋白酶体亚基可以改善蛋白质稳态并延长寿命。在小鼠中,通过在造血干细胞的转基因LAMP2a表达刺激CMA,可以提高目标细胞群的存活率,这与CMA的药理学增强可以缓解阿尔茨海默症和动脉硬化的结果相一致。因此,CMA的激活可能是延缓衰老的有效策略。
一项III期临床试验显示,在最近被诊断为ALS的患者中,使用降压药guanabenz可抑制病情向危及生命的延髓期的进展。Guanabenz可能会刺激真核生物翻译起始因子2α (eIF2α)的磷酸化(或抑制去磷酸化),这发生在作为UPR一部分的“整合应激反应(ISR)”中,尽管guanabenz的作用在多大程度上是由ISR的刺激介导的仍存在争议。重要的是,eIF2α的磷酸化RNA翻译从5’帽依赖的转变为非5’帽依赖的,一些延缓衰老的操作可以增强后者。此外,eIF2α磷酸化对于应激颗粒的诱导是必不可少的,应激颗粒对于限制饮食延长线虫寿命是必需的。最后,eIF2α磷酸化对于诱导自噬是不可或缺的,这是一种主要的延缓衰老机制,这表明UPR和自噬在延长寿命通路中存在串扰。未来的研究必须确定guanabenz减弱神经退行性病变的能力是否通过ISR刺激或替代机制介导。事实上,有人提出,ISR抑制剂也可以用于神经退行性疾病的治疗。
拓展:Nature重磅发现:衰老的根源在核糖体?衰老加剧核糖体暂停,破坏蛋白质稳态
五、自噬障碍
巨自噬(我们将称为“自噬”)包括将细胞质物质隔离在双膜小泡中,即自噬体,自噬体与溶酶体融合以消化腔内的内容物。因此,自噬不仅参与蛋白稳态,还可以影响非蛋白大分子(如异位胞质DNA、脂质囊泡和糖原)和整个细胞器(包括因“线粒体自噬”而功能失调的线粒体,以及其他导致“溶性自噬”、“网状自噬”或“前自噬”的细胞器),以及入侵病原体(“异种”)。衰老相关的自噬行为减少是细胞器更新减缓的重要机制之一,证明其可以作为衰老的新标志。需要注意的是,参与自噬过程的基因和蛋白质也参与了其他降解过程,如LC3相关的细胞外物质的吞噬以及细胞内废物(例如,功能失调的线粒体)排出,其随后被巨噬细胞清除。也就是说,有强有力的证据表明,自噬的核心过程与衰老有关(图3)。
自噬抑制加速衰老
在人类中,自噬相关基因如ATG5、ATG7, 以及BECN1的表达都随着年龄增长而下调。与年龄相近的对照组细胞相比,从长寿老人的后代中分离出来的CD4+淋巴细胞显示出更强的自噬活性。来自老年供体的B淋巴细胞和T淋巴细胞的自噬减少的同时,促自噬代谢物亚精胺的含量也下降。同样,在啮齿类动物中,一些细胞的自噬行为随着衰老逐渐退化,这一发现支持自噬随着年龄而变化的观点。自噬行为的减少可能导致蛋白质聚集物和失活细胞器的积累,抑制细胞对病原体的消除作用,并且增强炎症反应,因为自噬清除了炎症小体及上游的蛋白质。
自噬的基因抑制加速了模式生物的衰老。这一过程是部分可逆的,在小鼠中,Atg5在雷帕霉素诱导的shRNA影响下表达下调。Atg5的敲除会导致多器官系统的过早退化和衰老,从而导致个体过早死亡。停用强力霉素后,自噬行为恢复的同时伴随着全身炎症减弱,并在一定程度上延缓衰老。值得注意的是,在这个模型中,短暂的抑制自噬导致恶性肿瘤的发病率大幅增加。因此,推测自噬具有抑制肿瘤的作用,可能涉及细胞自主过程和癌症免疫监测。在患者中,调节或执行自噬的基因功能丧失可能导致心血管疾病、感染、神经退行性疾病、代谢疾病、肌肉骨骼疾病、眼部和肺部疾病,其中许多疾病在组织病理学和功能水平上类似于早衰。
拓展:Science:从小鼠到人类:雷帕霉素潜在的抗衰老功能
自噬刺激延缓衰老
有充分的证据表明,刺激自噬可以增加模式生物的寿命(表1)。过表达Atg5可以延长小鼠寿命、改善代谢和运动功能。此外,敲入突变的beclin 1(Becn1F121A/F121A)以减少Bcl-2的抑制作用,可以使自噬能力增强,达到延寿的目的。这种结果与衰老相关疾病和自发性肿瘤发生的减少,以及增加神经发生有关。
拓展:中科院昆明动物所孔庆鹏团队发现新型健康衰老促进因子ATF7
小鼠口服亚精胺可诱导多种细胞自噬,可延长约25%的寿命,同时延缓心脏衰老。后一种效应在心肌细胞特异性敲除Atg7时消失,这表明其延缓衰老的作用依赖于自噬。从机制上讲,亚精胺的促自噬作用与乙酰转移酶EP300的抑制作用(导致几个核心自噬蛋白的乙酰化减少)或与eIF5A的羟腐胺赖氨酸羟化作用有关,这对于自噬转录因子TFEB的合成有重要意义。在这些因素中,EP300是延寿药物去甲二氢愈创木酸和水杨酸的靶标。用C646抑制EP300可以模拟亚精胺对自噬和癌症免疫监测的激活作用。当老年循环B淋巴细胞或CD8+ T细胞在含亚精胺的条件下培养时,这些细胞的TFEB和eIF5A水平恢复到青少年时期,并伴有自噬能力的恢复。小鼠T细胞中的脱氧羟丁赖氨酸合成酶缺乏可以引发严重的肠道炎症,并伴有三羧酸循环的表观遗传重塑和重新连接,而用亚精胺处理野生型小鼠可预防结肠炎和结肠癌。因此,EP300抑制和eIF5A赖氨酸修饰可能都是亚精胺在体内作用的靶点。
拓展:
JAMA Netw Open:补充亚精胺对主观认知下降的老年人的认知和生物标志物的影响
Science:科学背书!从精液中发现的亚精胺,竟然有着抗衰老、抗癌、保护心血管和神经、改善肥胖和2型糖尿病等逆天神效
诱导线粒体自噬并可以延长小鼠寿命的药物包括NAD+前体(如烟酰胺、烟酰胺单核苷酸和烟酰胺核糖苷)和尿石素A。临床试验已经证明了NAD+前体在预防非黑色素瘤皮肤癌、降低糖尿病患者的胰岛素抗性,以及减少帕金森病患者的神经炎症等方面的疗效。此外,一项III期临床试验揭示了尿石素A增强肌肉力量和减少C反应蛋白(CRP)的能力。
拓展:
Nature重磅综述 | 深度解析代谢与细胞衰老间的恶性循环
eLife封面:吕宇轩博士发现明星药物雷帕霉素延缓衰老的新靶点
Front Endocrinol:上海第一人民医院周永团队完成大规模中国人群NAD+与衰老关系研究
六、营养感知失衡
营养感知网络在进化过程中是高度保守的。它包括细胞外配体,如胰岛素和IGFs,与它们相互作用的受体酪氨酸激酶,以及细胞内信号级联。这些级联反应涉及PI3K-AKT和Ras-MEK-ERK通路,以及转录因子,包括FOXOs和E26因子,它们可以反式激活参与各种细胞活动的基因。MTOR复合物1(MTORC1)的靶点能对包括葡萄糖和氨基酸在内的营养物质产生反应,也可以对缺氧和低能量等压力源产生反应,以调节转录因子的活性,如SREBP和TFEB。该网络是细胞活动的核心调节器,包括自噬、mRNA和核糖体产生、蛋白质合成、葡萄糖、核苷酸和脂质代谢、线粒体发生和蛋白酶体活动。营养感知网络通过以下方式对营养和压力状态做出反应,如果营养物质充足且压力水平较低,该网络通过激活合成代谢来利用营养,也可以通过诱导细胞防御途径来应对外来压力和营养短缺。营养感知网络与其他细胞内信号通路不仅可以互相影响,还存在广泛的网络内反馈。在不同的动物模型中,组成营养感知网络的基因活性降低可以增加寿命和改善健康(表1)。此外,对人类的遗传研究还涉及了FOXO3转录因子,以及影响人类寿命的营养感知网络的基因变体。表观遗传年龄也与人类细胞的营养感知有关。在青年时期,此网络的活动促进了有益的合成代谢过程,但在成年期,它开始表现促进衰老的特性(图4)。
生长营养轴是第一个已知的可调控衰老的轴,它是一个生长刺激级联反应,在其顶端,是由垂体产生的生长激素(GH)。生长激素作用于肝细胞的生长激素受体,刺激IGFs的分泌,特别是IGF1的分泌。IGF1通过IGF1R促进生长发育,通过激活PI3K-AKT和MTORC1网络来刺激营养信号。在多种模式生物中,该途径的自发或人为突变可以延长寿命,并缓解伴随衰老的功能退化(表1)。生长营养轴的先天缺陷会导致侏儒症,但从成年早期开始抑制这一轴会对机体健康产生有益的影响(图4)。
另一种参与营养感知的信号通路依赖于受体酪氨酸激酶ALK(图4),在小鼠中,摄食可以促进ALK在下丘脑中的表达,并由配体augmentor ɑ and β (Augɑ and Augβ)激活。在果蝇中,ALK的敲低可以降低甘油三酯水平和一些胰岛素样肽的表达,而通过基因或药物手段抑制ALK可以延长寿命,在雌性小鼠中表现更明显。在小鼠中,全身或下丘脑特异性的ALK缺失,以及Augα和Augβ的双敲除,都可以增强对肥胖的抵抗作用,而在人类中,ALK的失活突变也与减重有关。因此,该途径可能为干预代谢衰老提供额外的靶点。
针对癌症和代谢性疾病等疾病的药物往往与营养感知网络有关,因此,这类药物可以作为延缓衰老的候选药物。雷帕霉素和雷帕洛格,可以抑制MTORC1复合物,已经证明即使在成年后期开始治疗,也可以延长模式生物的寿命。在小鼠中,尽管雷帕霉素加剧了一些与衰老相关的特征,如白内障,但是它可以在其他方面改善健康,而且它对神经退行性疾病和其他与衰老相关疾病具有一定的治疗作用。
老年人易受病毒性呼吸道感染。MTORC1抑制剂的预治疗增加了老年志愿者对流感的免疫应答,并在随后的冬季减少了病毒性呼吸道感染,因此发现了可能逆转免疫衰老的策略。
拓展:
Science Advances:哈佛医学院科学家发现在生命早期服用雷帕霉素可延长10%的寿命
Aging (Albany NY).:科学家找到了“神药” 二甲双胍和雷帕霉素的“天然类似物”
机制
在人类中,IGF1在生命的第二个十年达到顶峰,但随着年龄的增长而下降。在成年或成年后期抑制GH/IGF1通路可以延长小鼠等模式生物的寿命。通过表达PI3K的阴性亚型p110α来抑制心脏IGF1R,可增加雄性小鼠的最大寿命,并改善老年小鼠的心脏功能。此外,酪氨酸激酶抑制剂可以抑制IGF1R酶,以此来改善肿瘤细胞中需要诱导自噬的免疫监测机制。长期使用IGF1R抗体可以提高雌性(但不是雄性)小鼠的寿命,同时减少炎症和肿瘤的发生。这些发现表明,IGF1/IGF1R信号轴可能成为延缓衰老的一个靶点。同时,在老年妇女(不低于95岁)以及老年综合人群(平均年龄76岁)中,低IGF1水平与认知障碍和低死亡率相关,这也支持上述猜想。此外,在英国生物样本库中,高IGF1与衰老相关疾病如痴呆、糖尿病、血管疾病、骨质疏松以及总死亡率之间存在显著正相关关系。在百岁老人中,IGF1BP2和IGFBP6的含量上升。未来研究将探索,尚未开发的抗体或小分子是否可以在不影响其他受体酪氨酸激酶(特别是胰岛素受体)的情况下选择性地抑制IGF1R信号传导,从而实现对生长激素轴的脉动抑制,进而实现副作用可控的健康益处。
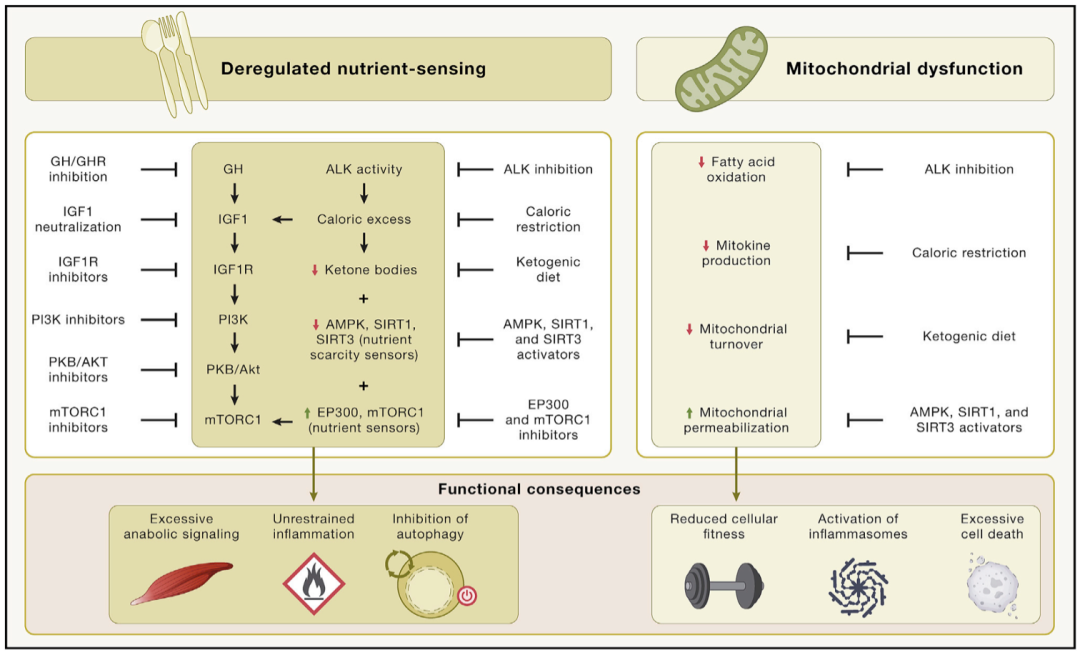
图4 代谢改变。这是交叉营养途径的简化版本,其展示了解除限制的营养感知过程及恢复营养感知的方法。值得注意的是,如果营养感知网络活性降低,衰老过程中除代谢外的许多过程都会受到影响,包括对不同应激源的反应、修复机制的激活、自噬激活和炎症控制。类似地,对于线粒体功能障碍,文章列出了一系列衰老相关的改变和可能有效的药物。衰老相关代谢改变引起的功能后果,其中一些与衰老的其他特征有关,在图表的下部举例说明。
营养影响
饮食是干预人类衰老最实际的靶标之一。从机制上讲,营养过剩:(1)触发细胞内的营养感应器,如mTORC1(由亮氨酸和其他氨基酸激活)和乙酰基转移酶EP300(由乙酰辅酶A激活);(2)抑制检测营养缺乏的感应器,如AMPK和乙酰转移酶SIRT1和SIRT3(对NAD+反应);以及(3)抑制分解代谢反应(糖原分解、与糖异生相关的蛋白分解和与酮生成相关的脂肪分解),从而抑制适应性细胞应激反应,包括自噬、抗氧化防御和DNA修复。相反,禁食和饮食限制会抑制mTORC1和EP300;激活AMPK、SIRT1和SIRT3;并刺激适应性细胞应激反应,因为它们抑制了生长营养轴,延长了包括灵长类在内的多种模式生物的寿命。
营养传感器是潜在的长寿药物的靶标(图4),饮食限制也可能益于健康和延长寿命。从机制上讲,这可以通过减少总的卡路里摄入量、控制饮食组成或限时饮食来实现。有实验发现雄性C57BL/6J小鼠在白天完全没有摄入营养,其生命长度则会得到延长,证明饮食限制方案在延长寿命方面有作用。但是,饮食限制方案并不能延长所有小鼠的寿命,这支持了该方案必须适应每个个体的基因构成的论点。在人类中,基于饮食限制的临床测试因依从性差而变得复杂,但对免疫和炎症有积极作用。
在临床试验中,间歇禁食(例如,禁食1天,然后自由进食1天)可以避免由热量限制引起的长期体重减轻,目前在小鼠中表现为延长寿命,以及在临床试验中改善与健康有关的生物标志物。类似的间歇禁食方案在果蝇中的寿命延长被归因于自噬刺激基因在夜间的特异上调,但这尚未在哺乳动物中进行研究。雷帕霉素诱导的寿命延长(在果蝇中部分依赖于自噬诱导)可以通过持续长期以及间歇性方案获得,这表明脉冲性抑制这一信号轴足以获得延长寿命的好处。临床上这种间歇治疗的最佳间隔尚未确定,不过每3-4周进行4-7天的部分热量限制可能足以改善代谢综合征和抗癌免疫监测。
拓展:
Cell子刊:间歇性禁食改变全身基因表达,延寿、降低疾病风险
Immunity:耶鲁大学研究证实,热量限制饮食能减少衰老相关蛋白,有助延长健康寿命
Nature Communications:科学家发现饮食限制延长寿命的机制及其关键成分
热量限制或可效延缓免疫衰老!科学家发现,热量限制可塑造肠道菌群,调节免疫细胞类型和比例,从而延缓免疫衰老
另一种可能有益的饮食是生酮饮食,这是一种低碳水化合物、高脂肪和充足蛋白质的饮食。禁食和生酮饮食都会增加酮体(特别是3-羟丁酸)的产生,酮体是由肝脏中的乙酰辅酶A以自噬依赖的方式合成的,在血浆中可以达到毫摩尔浓度,并取代葡萄糖作为维持大脑功能的基本燃料。在饮用水中长期但不是循环地服用3-羟丁酸可以延长小鼠的寿命和健康寿命。表明这种酮体介导了生酮饮食的一些有益效果。从机制上讲,3-羟丁酸可诱导血管扩张并激活作用于GTP蛋白偶联受体109A的免疫反应,而它直接抑制NLRP3,表明其潜在的多效性作用模式。
拓展:Aging Cell:只需2个月的生酮饮食,就足以改善大脑认知功能
七、线粒体功能障碍
线粒体不仅是细胞的动力源,也是炎症(当活性氧[ROS]或mtDNA从细胞器中泄漏并分别导致炎症小体或胞浆DNA传感器激活时)和细胞死亡(当半胱天冬酶、核酸酶或其他致命酶的激活剂从膜间隙释放出来时)的潜在触发因素。随着年龄的增长,线粒体功能因多种相互交织的机制而恶化,包括mtDNA突变的积累、蛋白稳态失衡导致呼吸链复合体的不稳定性、细胞器的周转减少、线粒体动力学的变化。这种情况降低了线粒体对细胞生物能量的贡献度,增加了ROS的产生,并可能触发导致炎症和细胞死亡的线粒体膜的透化。从逻辑上讲,线粒体的功能是维持健康,其逐渐退化会导致衰老表型(图4)。
线粒体功能与长寿
延长健康寿命的干预措施可能会刺激线粒体功能。例如左旋肉碱能够通过线粒体限制脂肪酸氧化,安慰剂对照试验显示对虚弱前期受试者和老年男性补充左旋肉碱都有积极影响(表1),这种作用可能是通过抵消与年龄相关的左旋肉碱水平的下降来实现的。但是与预期不同的是,在模式生物中损害线粒体的功能(线粒体低毒兴奋效应)能够延长寿命,前提是这种抑制是局部的并且发生在发育早期。在线虫中,部分抑制线粒体蛋白质的合成或输入是通过涉及线粒体UPR(UPRmt)的机制来延长寿命。在果蝇中,肌肉中特异性地敲除复合体I NDUFS1/ND75亚单位后会以UPRmt依赖的方式延长寿命。使用TPP-噻唑轻度抑制线粒体ATP的合成可以改善衰老小鼠的代谢健康,减少内脏脂肪,改善葡萄糖耐量、线粒体质量和氧化代谢。通过控制释放线粒体质子载体(CRMP)的方式部分解偶联肝脏线粒体也可以逆转高脂饮食诱导的肥胖小鼠的年龄相关代谢综合征。在非人类灵长类动物模型中,包括自发性肥胖的恒河猴和高脂、高果糖喂养的食蟹猴中,CRMP可以逆转代谢综合征的表现并改善脂肪酸氧化。这些作用与肝脏乙酰辅酶A水平降低有关,这是一种已知的刺激自噬的现象。CRMP诱导有丝分裂,这可能也解释了它们对新陈代谢的积极影响。二甲双胍,一种复合体I弱抑制剂,是一种抗糖尿病药物,已经作为延缓衰老的药物被人们广泛讨论。然而到目前为止,还没有证据表明通过调节线粒体可以延长人类的健康寿命或寿命。
拓展:Aging Cell:再添新证据,我国学者发现二甲双胍延长寿命新机制
由于缺乏血清/糖皮质激素调节激酶-1,线粒体膜通透性(MMP)增加从而减少了寿命,当自噬增强时,线粒体膜通透性进一步增加,但当自噬因基本的自噬相关基因敲除而被抑制时,线粒体膜通透性恢复正常。因此,线粒体膜通透性可能构成威胁生命的情况,自噬加剧了这种情况。已经开发出一种针对线粒体内膜双磷脂酰甘油的修饰后的四肽,Elamipretie,其与线粒体内膜蛋白腺嘌呤-核苷酸转运体-1结合以抑制线粒体膜通透性转变,这是导致MMP的一个特殊机制。Elamipreide对小鼠多种与衰老相关的表型具有积极作用,并在Barth综合征患者的临床试验中取得了积极的结果(表1)。Elamipretie是否可以与包括自噬增强剂在内的其他延长寿命的药物联合使用并产生有利效果,这一点很重要。除了这些研究进展,还有几项临床前研究和临床研究评估抗氧化剂亲脂阳离子MitoQ和SkQ1的潜在益处。未来的研究将进一步确定这些化合物在与其他干预措施联合的情况下,对改善衰老相关的线粒体功能障碍的效用。
线粒体微蛋白与衰老
血浆中线粒体DNA编码的小分子蛋白humanin的水平随着衰老而下降。然而,百岁老人及其后代表现出高水平的小分子蛋白。值得注意的是,humanin的水平与IGF1负相关,用GH或IGF1治疗GH功能不全的患者会降低humanin水平。humanin在线虫中的转基因表达通过诱导自噬延长寿命,humanin类似物HNG治疗中年小鼠可改善代谢健康并减少全身炎症。另一种mtDNA编码的小分子蛋白MOTS-c随着年龄的增加而下降,但可以通过运动诱导。MOTS-c促进代谢物AICAR的产生,其作为内源性AMPK激动剂,从而防止年龄依赖和高脂饮食诱导的胰岛素抵抗,以及饮食诱导的肥胖。因此,线粒体微蛋白作为潜在的延缓衰老因子,将细胞器功能与生物动态平衡联系起来。
拓展:Aging: 线粒体多肽humanin可以调节寿命和健康
八、细胞衰老
细胞衰老是一种由急性或慢性损伤引起的反应。衰老细胞在多种组织中的累积速度不同。和年轻人相比,老年人组织中衰老细胞累积到2-20倍,其中主要影响成纤维细胞、内皮细胞和免疫细胞。尽管在衰老过程中所有类型的细胞都会经历细胞衰老,这一过程在某种程度上是由端粒随年龄增长而缩短引发的。事实上,即使大脑或心脏等缓慢增殖的组织也可能含有衰老细胞。此外,衰老细胞在许多疾病中出现局部或特定组织的积累。通过遗传学或药理学手段持续消除衰老细胞延长了自然衰老小鼠的健康寿命,这是细胞衰老在衰老中的因果作用最令人信服的证据。此外,消除衰老细胞对许多以小鼠为模型的疾病具有治疗作用,目前至少已完成3项临床试验,15项临床试验正在进行。
引发原发性衰老的损伤类型包括癌基因信号、遗传毒性损伤、端粒过短、线粒体损伤、病毒或细菌感染、氧化损伤、营养失衡和机械应激。此外,继发性或旁分泌衰老可由细胞外炎症和纤维化介质包括CCL2、IL-1β、IL-6、IL-8和TGF-β触发。有证据表明原发性衰老和继发性衰老在生物学方面有所不同,但这种区别的分子基础仍不清楚。目前为止,细胞衰老最显著的特征是稳定的增殖停滞,可能由肿瘤抑制基因TP53和CDKN2A/p16及其下游效应蛋白CDKN1A/p21,以及RB1家族蛋白的激活所介导。这些蛋白共同抑制驱动细胞周期的CDK和转录激活因子(E2F家族)。衰老过程中的另一个重要事件是核纤层蛋白B1(Lamin B1)的耗尽。这导致Lamin相关的异染色质的丢失和富含H3K9me3的异染色质的重新形成,这一过程可通过HP1α或SAHFs(Senescence Associated Heterochromatin Foci)观察到,最终结果是长期的增殖停滞和低自发逃逸率。根据它们的分子组成,暴露于基因毒性治疗的癌细胞可能会经历具有高度稳定的细胞周期阻滞的典型衰老反应,或者可以经历具有高度可逆的类似衰老反应,甚至可以完全绕过衰老。值得注意的是,在胚胎发生期间,衰老也在特定细胞和结构的程序性消除中发挥作用。
拓展:Nature子刊:抗衰老疫苗来了?打一针,清除衰老细胞,延长寿命
衰老和人类疾病
细胞衰老与多种非传染性疾病有关,包括肺纤维化、肾脏疾病、肝脂肪变性、肥胖相关代谢综合征、I型和II型糖尿病、动脉粥样硬化、阿尔茨海默症和帕金森症。细胞衰老在这些疾病中的致病性可以通过衰老相关分泌表型(SASP)来解释。SASP源于衰老细胞的三个特征:(1)内源逆转录病毒的转录去抑制,尤其是LINE-1,导致双链DNA泄漏到胞浆,进而激活cGAS/STING和TLR通路;(2)线粒体ROS的大量产生;(3)自噬-溶酶体系统的扰动导致溶酶体含量增加,辅助溶酶体衰老相关β-半乳糖苷酶(SABG)的组化检测。
SASP是高度异质性的,依赖于先天免疫信号通路(cGAS/STING、TLRs和NLRPs)、mTORC1和转录因子(NF-kB、CBPs、GATA4等)的细胞类型特异性激活。SASP通常同时对微环境有部分矛盾的影响:(1)通过分泌趋化因子(CCL2、CXCL2和CXCL3)和细胞因子(IL-1β、IL-2、IL-6和IL-8)招募和激活免疫细胞;(2)通过分泌TGF-β抑制免疫系统;(3)通过促纤维化因子(TGF-β、IL-11和PAI1)激活成纤维细胞活化和胶原沉积;(4)通过分泌基质金属蛋白酶重塑ECM;(5)通过分泌生长因子(EGF和PDGF)促进祖细胞的激活和增殖;(6)触发邻近细胞的旁分泌衰老(TGF-β,TNF-α和IL-8)。在多种疾病中,SASP的净效应是导致慢性炎症和促进纤维化。
虽然没有单一明确的细胞衰老marker,但这一过程可以通过共同存在的特征组合来识别,这些特征组合是特异性的,并为这些特征组合提供了分子定义:(1)由SABG检测的溶酶体扩张;(2)CDK抑制剂上调,特别是p16和/或p21;(3)核膜中LMNB1蛋白丢失;(4)染色质成分HMGB1从细胞核中丢失,并作为一种预警蛋白在细胞外释放;(5)异染色质灶,显像为HP1γ灶或SAHFs灶;(6)ROS水平升高;(7)DNA损伤加重,可见γH2AX灶;(8)高水平的SASP因子,特别是IL-6、TGF-β、PAI1等。
鉴于细胞衰老与多种病理之间的联系,目前提出了关于这种细胞反应的生物学目的的问题。细胞衰老是一种强有力的肿瘤抑制机制,但越来越多的证据表明,细胞衰老与组织修复过程有关,在这个过程中,衰老细胞促进局部纤维化,并募集免疫细胞,清除受损和衰老细胞。在这方面,组织修复被认为是一个两步走的过程:细胞衰老,然后是免疫细胞招募和免疫清除衰老细胞(图5A)。在这种情况下,衰老是一种暂时的受限制反应,它的自我消除是一个有益结果。只有当第二步的免疫清除没有实现时,衰老的病理特征才变得明显,衰老细胞的积累和SASP对组织微环境的作用最终导致纤维化。
清除衰老细胞(Senolytics)
细胞衰老与多种病理之间的密切联系促使人们寻找选择性杀死衰老细胞的小化合物,即所谓的“延缓衰老类药物”。值得注意的是,senolysis(衰老细胞的清除)与衰老反应的清除有很大的不同,例如p16或p21的突变可以导致后者。senolysis不能阻止衰老的发生,而是重现了衰老细胞的天然免疫性清除(图5A)。为了支持这一观点,使用长期遗传诱导或药物诱导的衰老小鼠表现出寿命的延长,而没有增加癌症发病率或出现组织修复缺陷的现象。
拓展:抗衰老公司Altos Labs发表Nature论文:衰老细胞会终生损害身体,去除衰老细胞有助于抗衰老
目前senolytic疗法的数量仍然有限,但一些药物已广泛应用于疾病的临床前模型,如维奈托克、达沙替尼和槲皮素的联合治疗(D/Q)、非瑟酮、强心苷等。衰老细胞的存活和抗凋亡能力强烈依赖于BCL2蛋白家族,特别是BCLXL、BCL2和BCLW。这使得靶向这三种蛋白质的BCL-2抑制剂对清除衰老细胞非常有效。BCL-2抑制剂已在临床试验中评估了抗肿瘤活性,预计该药物(或对血小板无毒性的衍生物)将进入衰老相关疾病的临床试验。其它潜在的延缓衰老类药物,如D/Q和非瑟酮已被批准用于人体,并正在针对多种适应症进行各种临床试验,但它们的机制尚不清楚。达沙替尼是一种混杂激酶抑制剂,而槲皮素和非瑟酮是具有多靶点的天然类黄酮。D/Q已经在临床试验中进行了测试,对治疗肺和肾纤维化有很好的结果。强心苷抑制所有细胞中存在的细胞质膜Na+/K+-ATPase,导致阳离子失衡并降低细胞内pH值。强心苷的衰老溶解机制可能与衰老细胞对细胞内低pH值的脆弱性有关。因此,对谷氨酰胺酶的化学抑制使细胞丧失了抵抗低pH值的能力,从而导致细胞溶解。所有上述讨论的延缓衰老类化合物均在较多的衰老相关小鼠疾病模型上发挥治疗活性。senolysis也可以通过免疫方法实现,靶向出现在衰老细胞表面的蛋白能够改善小鼠衰老相关疾病模型,特别是通过靶向糖蛋白NMB(GPNMB)的抗体和靶向受体uPAR的CAR-T细胞。
综上所述,细胞衰老是对应激和损伤的重要反应,在正常生理学中,免疫清除紧随其后,但在衰老或慢性损伤时无法被免疫机制消除,因此由于促炎和促纤维化因子的大量分泌而致病。旨在清除衰老细胞的治疗策略已在动物模型中进行了广泛的探索,目前正在进行临床试验(表1)。
拓展:抗衰老药物Senolytics翻车?最新Science论文显示,其清除的“僵尸”细胞具有保护作用
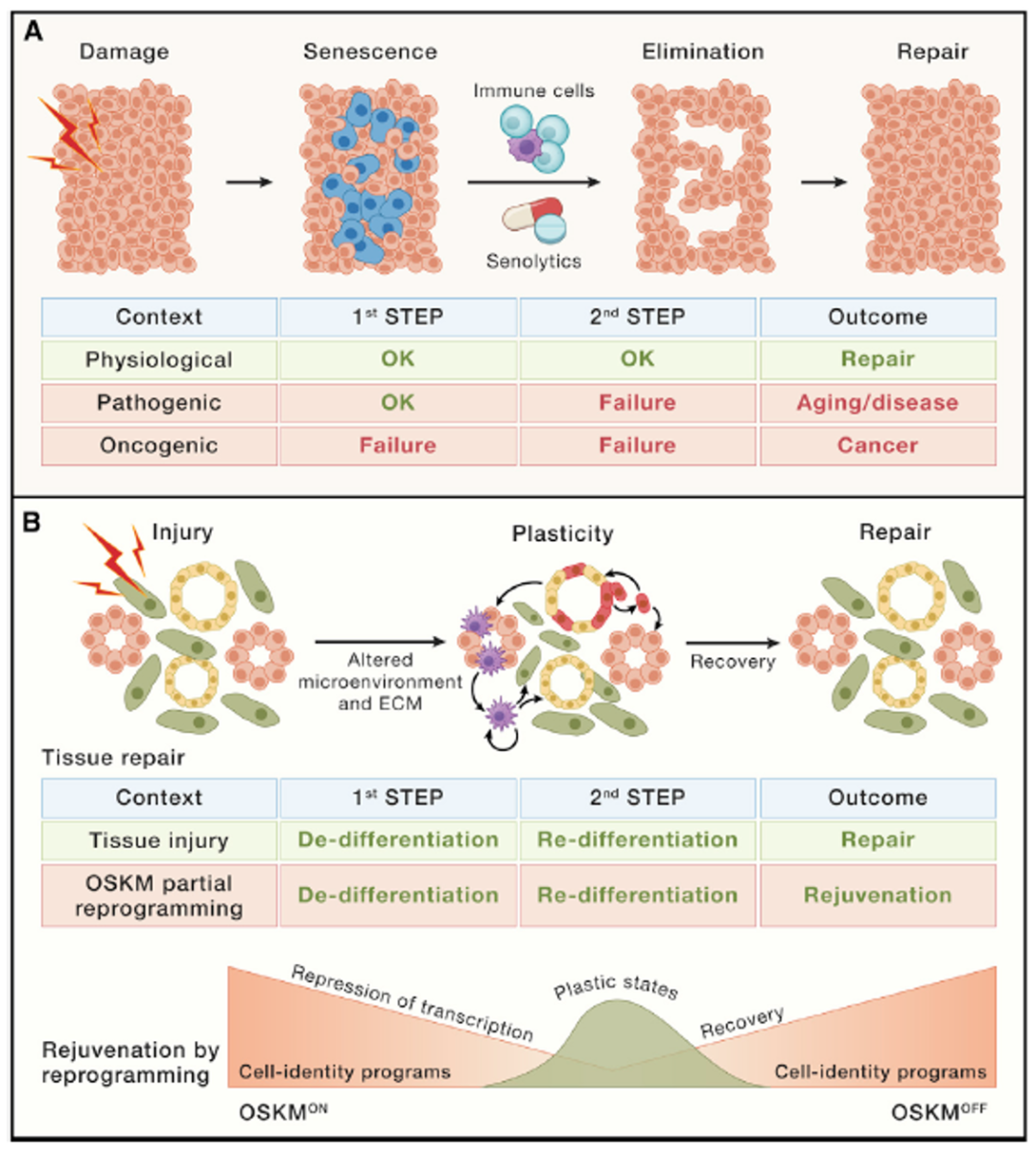
图5 细胞衰老和干细胞耗竭。(A)细胞衰老通常促进损伤后的组织修复,保护机体免受致癌损伤。这是通过两个步骤实现的:(1)建立衰老和(2)招募免疫细胞以消除衰老细胞,从而促进组织修复。如果这些步骤中的任何一步失败了,有机体就容易患上疾病。(B)干细胞耗竭是由于组织修复所需的细胞可塑性丧失造成的。组织修复需要通过分泌细胞因子(部分与于衰老相关的分泌反应相关)、生长因子和细胞外基质(ECM)的调节剂来改善微环境,这有利于来自不同组织区域细胞获得去分化和可塑性。这些损伤诱导的可塑性细胞可能获得多能干细胞功能。OSKM因子的瞬时表达抑制细胞身份程序的转录,导致全局去分化(OSKMon)和可塑性的获得。为了恢复活力,这个过程必须在这个点中断(OSKMoff),以允许细胞重新分化并恢复其原始细胞身份。
拓展:Science子刊:衰老细胞清除疗法,可增强器官移植成功率,减少并发症
九、干细胞耗竭
衰老与稳定状态下的组织更新衰退有关,也与损伤后的组织修复受损有关,每个器官都有自己的更新和修复策略。例如,在骨骼肌中,一种处于单能和单向层次结构顶端的卫星细胞,用于组织更新和修复。在暴露于损伤处和高更新频率的皮肤表皮,有多个干细胞niche,特别是在毛囊,每个细胞产生自己的子细胞和占据自己的位置。然而在损伤后,多个细胞可以获得干细胞特性并破坏边界。肝脏、肺或胰腺等其它器官在正常情况下表现出较低的更新频率,与获得的干细胞特性(包括不同细胞类型的增殖和多能性)形成对比(图5B)。事实上,组织修复在很大程度上被认为依赖于损伤诱导的细胞去分化和可塑性。例如,在肠、脑和肺中,损伤诱导非干细胞去分化,从而重新激活通常沉默的干性转录程序,从而获得组织修复所需的可塑性。在正常的稳态条件下,损伤诱导的可塑性(随年龄增长逐渐丧失)可能比常驻干细胞的可塑性更与衰老相关。干细胞和祖细胞与没有干细胞潜能的细胞一样,都具有相同的衰老特征,因此本文在这里不讨论关于衰老的每个特征对干细胞功能影响的大量文献。相反,关注于一种基于“细胞重编程”概念的对抗干细胞功能随衰老而衰退的策略。这个过程被认为以细胞自主的方式作用于多种细胞类型;然而,由于它的长期影响,其对干细胞和祖细胞的影响被认为具有更高的相关性。
Cell Discovery:刘光慧等团队发现抗衰老关键代谢物——尿苷
通过重编程实现组织修复再生
细胞向多能性的重编程包括通过四种外部转导的转录因子,即OCT4、SOX2、KLF4和MYC(OSKM),将成体体细胞转化为胚胎多能细胞(被称为诱导性多能干细胞或iPSC)。重编程过程通常需要数周时间,在此期间,细胞首先通过细胞身份基因的转录抑制失去其分化表型,随后激活多能性基因。完全重编程不仅意味着细胞身份的改变,还意味着细胞再生,其特征在于许多衰老特征被重置为胚胎状态,如p16减少、端粒延长和DNA甲基化时钟的重置。有趣的是,在去分化开始后不久,这种恢复以渐进的方式发生。事实上,可以使用OSKM启动重编程,在中间状态中断过程,并允许细胞恢复其原始身份。这种短暂的细胞扰动,也被称为“部分”、“短暂”或“中间”重编程,能够在体内外使衰老的细胞标志物恢复到年轻化的状态,例如DNA甲基化时钟、DNA损伤、表观遗传模式和转录组中的衰老相关变化。因此,去分化和再生过程可以认为是耦合的。具体来说,去分化意味着表观遗传和转录程序的消除,这也可能消除与衰老相关的变化。部分重编程中断后,细胞在重新分化过程中重新建立其原始的表观遗传和转录状态,有趣的是,细胞不会重新建立已消除的衰老相关变化,因此将表观基因组和转录组重置为更年轻的状态。
小鼠的瞬时重编程赋予了老年组织修复能力,因此后续的损伤可以像在年轻个体中一样得到有效修复。这种增强的修复能力已经在胰腺(内分泌部)、骨骼肌、神经纤维、眼、皮肤、心脏和肝脏的组织损伤模型中得到证实。此外,自然衰老的组织功能障碍特征,如视力下降、海马体中神经发生和长期记忆的丧失,也可以通过瞬时重编程而部分逆转。在少数情况下,瞬时重编程在组织修复过程中也是有益的(而不仅仅是在受伤之前),创伤性脑损伤和皮肤伤口愈合就是这种情况。最后,应该提及的是,早衰小鼠的寿命可以通过瞬时重编程来延长,但尚未有野生型小鼠通过OSKM延长寿命的报道。
部分重编程重现了自然组织修复的特征(图5B)。在这两种情况下,细胞都会经历一个短暂的去分化过程,获得胚胎和祖细胞特征,随后再分化。因此,去分化和再分化可以解释组织再生,这与观察到的肌细胞的瞬时去分化,然后再分化,继而诱导转录组的年轻化现象一致。组织修复的自然过程可能意味着某种程度的细胞再生,这与表观遗传甲基化时钟在组织损伤后很快加速并在组织修复过程中部分逆转的发现一致。此外,据报道,组织损伤会产生一个组织微环境,该微环境非常适合IL-6驱动的重编程。最后,转录因子FOXM1的循环表达延长了早衰小鼠和野生型小鼠的寿命。虽然详细机制尚未探明,但FOXM1在肾脏损伤后被诱导,并在修复过程中参与触发肾小管上皮细胞的去分化和增殖。因此,自然组织修复和人工重编程的几个特征可能会趋于一致,或许可以改进恢复衰老组织修复能力的策略。
拓展:Front aging neurosci-衰老特征对iPSC衍生以及重编程神经元的影响
十、细胞间通讯的改变
衰老与细胞间通讯的逐渐改变有关,这会增加系统中的噪音并损害稳态和激素调节。因此,衰老会伴随神经、神经内分泌和激素信号通路的缺陷,包括肾上腺素能、多巴胺能和基于胰岛素/IGF1和肾素-血管紧张素的系统,以及与生殖功能丧失相关的性激素。虽然在SASP中清楚地表明这种改变的主要原因源于细胞内,但细胞间通讯的紊乱最终总结为一个标志,将细胞内在标志与系统性细胞标志联系起来,包括炎症反应的长期化、对病原体和癌前细胞的免疫监视能力下降,以及人类基因组和微生物组之间双向通讯的改变最终导致生态失调。这方面的许多研究都集中在寻找具有促衰老或延长寿命特性的血源性系统因素、细胞间不同通讯系统的作用,以及对衰老过程中ECM破坏的功能相关性的评估。
促衰老的血源性因子
单次输注老年血液可在数日内诱发年轻小鼠衰老,而用含5%白蛋白的生理盐水缓冲液简单稀释老年小鼠的血液可诱发多种组织年轻化,这说明存在加速衰老进程的循环因子。在促衰老血源性因子中,趋化因子CCL11/eotaxin和炎症相关蛋白β2-微球蛋白抑制神经发生,IL-6和TGF-β损害造血功能,补体因子C1q损害肌肉修复。从理论上讲,中和这些因素可能具有强大的延缓衰老作用。事实上,上述因素中的几个是在SASP的背景下分泌的,并且可能是“传染性”衰老现象的共同原因,这也涉及细胞外囊泡。因此,所谓的“senomorphics” (又被称为SASP抑制剂和Senostatics)可能被用来抑制SASP和延缓衰老。
拓展:
Science Advances:抗衰老新方法:注射年轻个体的细胞外囊泡,改善衰老迹象
Nature Metabolism:“换血抗衰老”奠基人最新研究:老年血液会迅速诱导年轻个体未老先衰
延缓衰老的血源性因子
年轻小鼠血液中存在的可溶性因子可有效恢复老年小鼠的更新和修复能力(表1)。异体共生实验和广泛的单细胞转录组学研究证实了年轻血液能够使多个组织恢复活力,并恢复与衰老相关的下调表达基因,特别是参与电子传递链的线粒体基因。趋化因子CCL3/MIP-1α作为造血干细胞和祖细胞的再生因子;金属蛋白酶抑制剂TIMP2与海马体再生相关;抗炎白细胞介素IL-37(在老年人的单核细胞中减少)提高了老年小鼠耐力运动并改善其全身新陈代谢;细胞因子GDF11能使一些组织(肌肉、大脑和胰腺内分泌部)恢复活力,尽管它由于其促纤维化的副作用而损害其他组织的功能和修复;遗传增强的VEGF过表达小鼠表现出肝脏和肌肉修复能力增强,整体健康状况改善,平均寿命延长约40%。
拓展:
Nature重磅:斯坦福科学家首次在人脐带血中发现改善大脑衰老的蛋白!
远程和短程通讯系统
中枢神经系统控制影响周围器官衰老的多个方面,这解释了大脑特异性基因操作(如过表达SIRT1、UCP1,或敲除IKBKB和TRPV1)如何延长小鼠寿命(表1)。这些远程活动的确切机制尚待确定。值得注意的是,细胞间通讯还涉及细胞外分子(如ROS、一氧化氮、核酸、前列腺素和其他亲脂性分子)、从白色脂肪组织(adipokines)、棕色脂肪组织(baptokines)、心脏(cardiokines)、肝脏(hepatokines)和骨骼肌(myokines,包括运动时产生的“运动因子”)等各种组织中释放的可溶性因子、细胞结合的配体和其他细胞上的受体(如可以维持细胞结合的IL-1α)之间的互作,以及由紧密连接或间隙连接介导的细胞间直接相互作用。所有这些通讯系统在衰老过程中都可能发生变化,因此其潜在的促进和延缓衰老的特性正被细致研究。
细胞外基质
衰老会对ECM的长效蛋白质成分造成大量损害,包括AGEs、羰基化和氨基甲酰化、弹性蛋白断裂和胶原蛋白交联,从而导致组织纤维化(纤维化衰老)。这一有害过程部分是由于过度释放TGF-β和其他生长因子,以及转录因子TAZ和YAP的核转位,它们作为机械转导器,触发了促纤维化基因(如转谷氨酰胺酶-2、赖氨酰氧化酶(LOX)和LOX样酶)的表达。ECM刚性也会影响衰老细胞的功能,而衰老细胞又会分泌基质金属蛋白酶,放大ECM的损伤,并通过蛋白水解产生损伤相关的分子以激活促衰老、促纤维化和促炎症通路。衰老基质的刚性增加也可能有利于WNT信号诱导成纤维细胞的激活和促纤维化基因的表达。该通路与其他促纤维化通路(如NOTCH、RAS、TGF-β/SMAD和hedgehog/GLI)表现出广泛的串扰,从而证明了年龄相关的纤维化发展机制的复杂性和相互联系。值得注意的是,由基质刚性引起的机械变化足以导致少突胶质细胞祖细胞年龄相关的功能丧失,这一过程由机械响应性离子通道PIEZO1介导。
已有研究提供了ECM刚性对衰老影响的因果证据,并提出了改善健康衰老的方法(表1)。使用AAV载体对Piezo1进行体内抑制,可使老年小鼠大脑中的少突胶质细胞祖细胞恢复活力。基质细胞中YAP/TAZ的基因失活会加速衰老,尽管维持YAP功能可使衰老细胞恢复活力,并通过控制cGAS-STING信号传导防止衰老特征的出现。此外,可产生抗胶原酶I型胶原蛋白(Col1a1r/r)的转基因小鼠表现出血管细胞衰老、加速衰老和寿命缩短。胶原蛋白对人类长寿的重要性已经在COL25A1(编码大脑特有的胶原蛋白)中的罕见变异而得到加强,这种胶原蛋白可能对阿尔茨海默病有保护作用。此外,由年轻的人类成纤维细胞制备的ECM可诱导老年人的衰老细胞进入年轻状态。硫酸软骨素和透明质酸等ECM化合物可恢复与衰老相关的胶原蛋白下调,并延长线虫的寿命。相反,透明质酸酶TMEM2的异位表达可通过改变p38/ERK MAPK信号通路来促进对内质网应激的抵抗,并延长线虫的寿命。在小鼠中,软骨素6-磺基转移酶的缺失导致大脑中ECM异常、早期记忆丧失并加速大脑衰老,而过表达这种酶则能改善老年小鼠的记忆力。回顾性分析表明,口服氨基葡萄糖/硫酸软骨素可降低人类的全因死亡率。然而,到目前为止,还没有前瞻性证据表明这种延长寿命的效果可以通过ECM的改善来调节。
十一、慢性炎症
炎症在衰老过程中增加("炎性衰老"),伴有全身性表现以及病理性局部表型,包括动脉硬化、神经炎症、骨关节炎和椎间盘退化。相应地,炎性细胞因子和生物标志物(如CRP)的浓度随着年龄的增长而增加。血浆中IL-6水平的升高是老龄人群全因死亡率的预测性生物标志物。随着炎症加剧,免疫功能下降,这种现象可以通过对患者和小鼠组织血液中的髓系细胞和淋系细胞进行高维监测来捕捉。例如,与年龄相关的T细胞群(称为Taa细胞)由耗竭状态的记忆细胞组成,这些细胞通过颗粒酶介导促炎作用。T细胞群的变化引起促炎症的TH1和TH17细胞功能亢进,免疫监视功能缺陷(对消除病毒感染、恶性或衰老细胞有负面影响),自我耐受性丧失(随之而来的是与年龄相关的自身免疫性疾病的增加),以及生物屏障的维持和修复减少,这些都有利于系统性炎症(图6A)。
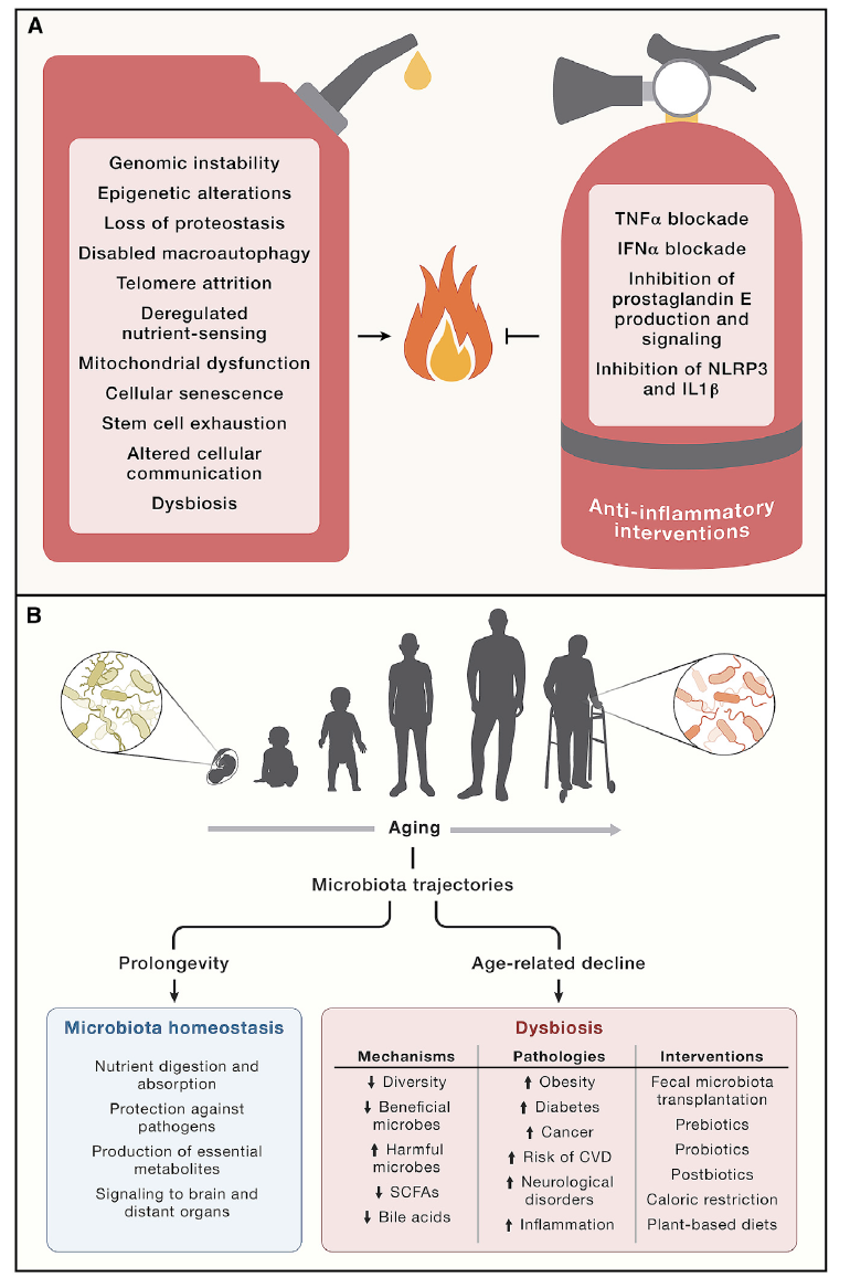
图6 细胞外功能紊乱。细胞间通讯的改变将细胞内在特征与包括慢性炎症在内的系统性细胞特征(包括慢性炎症),以及人类基因组和微生物组之间串扰的改变联系起来,最终导致生态失调。(A)衰老过程中的慢性炎症是源于其他所有特征引起的多种紊乱。图中右侧展示了几个具有代表性的对健康寿命和寿命有积极影响的抗炎干预措施的例子。(B)生态失调导致了与衰老相关的多种病理状况。人类肠道微生物群落在衰老过程中发生显著变化,最终导致了生态多样性普遍下降。右图下部展示了这些微生物群变化机制的主要特点,以及可以促进健康衰老的肠道微生物群组成的干预示例。CVDs,心血管疾病;SCFAs,短链脂肪酸。
拓展:一文读懂:炎症性衰老
炎症和其他衰老特征之间的联系
炎症是源于衰老其他的特征的多种紊乱导致的结果。例如,炎症可以由细胞核和线粒体DNA易位到胞浆中引发,特别是当自噬失效、无法拦截异位DNA时,在那里它们可以刺激促炎DNA感受器。基因组的不稳定性有利于不确定潜能的克隆造血(CHIP),通常伴随着具有促炎表型的髓系细胞的扩张,导致心血管老化。有趣的是,最常见的CHIP相关突变影响表观遗传修饰酶DNMT3(使DNA中的胞嘧啶残基甲基化)和TET2(催化甲基胞嘧啶氧化为5-羟甲基胞嘧啶)。从机理上讲,影响TET2的CHIP会增强髓系细胞产生IL-1β和IL-6,并促进心血管疾病(cardiovascular disease,CVD),这个效应在IL-6受体功能缺失突变或用IL-1β中和抗体治疗的个体中减弱。
促炎蛋白的过度表达可继发于表观遗传失调、蛋白稳态失衡或自噬功能受损。过度的营养信号导致GH/IGF1/PI3K/AKT/mTORC1轴的激活,从而引发炎症。此外,继发于衰老细胞积累的SASP以及细胞外碎片和感染性病原体的积累(由于衰老和髓系细胞、淋巴细胞的衰竭而无法清除)也会促进炎症。后一种现象涉及衰老相关的胸腺退化、胸腺生成的降低,从而导致T细胞库的减少,无法对新抗原产生有效的免疫应答。值得注意的是,人类的胸腺细胞生成可以通过CR改善,并且在小鼠中可以敲除CR下调基因PLA2G7来对抗胸腺萎缩。最后,昼夜节律紊乱和肠屏障功能障碍也会加剧炎症。
抗炎、延缓衰老干预手段
尽管全身炎症与上述所有衰老相关的改变有机制联系,但炎症本身就是一个衰老的标志。事实上,通过对炎症和免疫系统进行特定处理可以加速或延缓不同器官系统的衰老过程。例如,线粒体转录因子A(mitochondrial transcription factor A,TFAM)中T细胞特异性缺陷足以驱动心血管、认知、代谢和身体衰老,并伴随循环细胞因子增加。TNF-α抑制剂依那西普部分逆转了这种表型。小鼠造血细胞中DNA修复蛋白ERCC1的杂合缺失足以诱导非淋巴器官的免疫衰退和老化,以及许多与寿命缩短相关的器官损伤迹象,这种表型可以通过非瑟酮缓解。这些结果支持了免疫系统老化可能导致机体衰老的观点。值得注意的是,将TFAM缺失的T细胞、年轻ERCC缺陷型脾细胞或年老的野生型脾细胞过继转移到年轻小鼠中会诱导衰老,而将年轻免疫细胞移植到ERCC缺陷小鼠中可减缓衰老,这表明免疫细胞有调节机体衰老的能力。
抗炎治疗延长寿命例子有很多。因此,阻断TNF-α可防止小鼠的肌细胞减少,并改善衰老大鼠的认知能力。阻断常见的1型干扰素受体(type 1 interferon receptor,IFNAR1)可逆转衰老小鼠肺中单核细胞的积聚。敲除骨髓细胞中前列腺素E2受体EP2或用EP2药物抑制剂治疗老年小鼠可改善认知。敲除炎症小体蛋白NLRP3可改善代谢生物标志物、葡萄糖耐受、认知和运动能力,延长小鼠寿命。NLRP3或其下游酶胱冬肽酶-1的药理学抑制剂对正常和加速衰老模型具有令人鼓舞的临床前效应。最重要的是,卡那单抗对胱冬肽酶-1产物IL-1β的抑制是适用于患者的延缓衰老治疗的例证。第III期临床试验CANTOS评估了卡那单抗预防有心肌梗死病史和明显炎症迹象的患者复发CVD的能力。除了达到试验的主要终点外,卡那单抗还降低了糖尿病和高血压的发病率,以及肺癌的发病率和死亡率。最后,尽管长期使用非甾体抗炎药(如阿司匹林)可能对人体健康产生积极影响,特别是在预防CVD和胃肠道癌症方面,但一项大型的III期临床试验(对70岁以上的受试者服用阿司匹林)产生了负面结果。因此,有必要进行进一步的研究以探讨在年轻时使用阿司匹林进行预防性治疗的价值,或将阿司匹林与其他药物结合使用,或用毒性较小的抗炎药替代阿司匹林。
拓展:
无需换血,阻断IL-1信号,就能恢复衰老血液的年轻活力,逆转衰老
十二、肠道微生态失调
近年来,肠道微生物已成为多种生理过程中的一个关键因素,如营养消化和吸收、抵御病原体和生产必需代谢产物(例如维生素、氨基酸衍生物、次级胆汁酸和短链脂肪酸(short-chain fatty acids,SCFAs))。肠道微生物群还向外周/中枢神经系统和其他远处的器官发出信号,并对宿主健康的总体维持产生强烈的影响。细菌-宿主双向交流的扰乱导致了肠道生态失调,并导致多种病理状况,如肥胖、2型糖尿病、溃疡性结肠炎、神经系统疾病、CVD和癌症。这一领域的进展引起了人们对探索衰老过程中肠道微生物群变化的极大兴趣。
衰老过程中的微生物群变化
由于宿主遗传多样性(种族)、饮食因素、生活习惯(文化)以及地理环境条件,肠道内的微生物在个体之间高度多变,这使得人们难以揭示微生物群与多效性年龄相关疾病表现之间的关系。尽管如此,一些Meta分析揭示了微生物群与疾病的相关性,这些相关性已在不同的疾病和国家得到验证。对人类和动物模型的研究提供了有关临床、流行病学、社会学和分子基础方面(老年微生物组对人类健康和疾病的复杂影响)的宝贵信息。一旦在儿童时期建立了菌群的多样性,在成年时期其就保持相对稳定。然而,在衰老过程中菌群的结构和活性逐渐发生变化,最终导致多样性的总体下降。因此,对百岁老人进行的几项研究表明,丰度大的几个核心菌群(如拟杆菌属和罗氏菌属)减少了,但双歧杆菌属和阿克曼菌属等几个属的菌群增加了,这似乎具有延长寿命的作用。
人们通过对最近来自三个年龄跨度为18-101岁的独立队列的9000多名个体的肠道微生物组和表型数据的分析,拓展了这方面的研究。值得注意的是,随着年龄的增长,每个人的肠道微生物群变得越来越独特,这种独特性与参与免疫调节、炎症和衰老的微生物代谢产物有关。在老年人中,健康的个体表现出对独特微生物成分的持续偏移,而在健康状况较差的个体中,这种偏移减少或消失。健康衰老的微生物群模式的特征是核心类群的耗竭,如大多数人中存在的拟杆菌。此外,在接近极端年龄的个体中,拟杆菌高水平的保留和低肠道微生物组的独特性与生存率下降显著相关。然而,百岁老人和超百岁老人的微生物群研究结果并不总是与老年人群的结果一致。一项名为ELDERMET的研究发现,与年轻对照组相比,老年人的核心属拟杆菌属、另枝杆菌属和拟副杆菌属的优势增加。这项研究还确定了与虚弱、认知、抑郁和炎症相关的肠道微生物群组成的年龄相关性变化。另一项研究揭示了不同种族人群中的微生物群与年龄相关的轨迹,以及肠道微生物群中普遍的衰老相关的性别依赖性差异减少。值得注意的是,老年人中几种促进健康的细菌种类表现出的更高丰度,包括阿克曼菌。这些结果表明,除了老年人的饮食变化和生活方式外,与衰老相关的生理变化可能会对人体肠道微生物群产生深远影响。
所有这些研究结果的异质性表明,衰老可能存在多种肠道微生物组轨迹。然而,微生物群产生的氨基酸衍生物的血浆浓度有一个有趣的趋同。这些代谢产物包括由肠道细菌降解色氨酸产生的吲哚,以及苯丙氨酸/酪氨酸的几种发酵产物,如对甲酚硫酸盐、苯乙酰谷氨酰胺和对甲酚葡萄糖醛酸。这一发现与ELDERMET的数据一致,表明粪便中对甲酚的浓度与虚弱程度的增加相关,并可能导致该人群年龄相关的机能下降。相反,某些吲哚代谢产物的血浆浓度与老年人健康状况的改善相关。吲哚代谢产物至少部分地通过与芳香烃受体结合来减轻炎症反应,从而增加小鼠的健康期和寿命。
对百岁老人肠道微生物组的进一步代谢组学和功能分析表明,它富含一些特定的细菌,如Alistipes putredinis和Odoribacter splanchnicus。这些细菌中的一些能够产生独特的次级胆汁酸,包括异丙胆酸,它对艰难梭菌和粪肠球菌等革兰氏阳性多药耐药病原体具有强大的抗菌作用。因此,特定的胆汁酸代谢可能降低病理生物感染的风险,并有助于肠道内稳态,从而降低年龄相关慢性疾病的易感性。
拓展:Aging Cell:新思路,靶向肠道菌群或为治疗衰老及衰老相关疾病的可行方案
粪便菌群移植与衰老
病理性衰老的多组学研究表明,两种不同的早衰小鼠模型表现出肠道生物失调,其主要特征是变形杆菌和蓝细菌的丰度增加,以及疣状菌的水平降低。与这些发现一致的是,患有HGPS或NGPS的人类早衰患者也表现出肠道生物失调,而长寿的人表现出变形杆菌的显著减少和疣微菌的显著增加。粪便微生物群移植(Fecal microbiota transplantation,FMT)在体内证明了这些变化的因果关系。野生型小鼠的肠道微生物移植到两种早衰小鼠模型中能提高早衰小鼠的健康期和寿命,而给予嗜黏蛋白阿克曼菌Akkermansia muciniphila也足以达到这种效果。相反,从早衰供体到野生型小鼠的FMT诱导了有害的代谢改变。早衰小鼠体内次级胆汁酸和其他代谢产物的恢复表现出重建健康微生物群的有益效果。
拓展:Gut:Akk菌或能防肠癌!南京医科大团队发现,灭活的Akk菌或其外膜蛋白可以调节免疫响应,改善肠炎,预防肠癌
FMT还揭示了肠道生态失调在慢性全身炎症和与衰老相关疾病相关的适应性免疫下降中的致病作用。将肠道微生物群从老年小鼠转移到年轻无菌小鼠引发炎症反应,其特征是脾脏中CD4+ T细胞分化增强,炎性细胞因子上调,细菌来源的炎性因子增加。FMT也为肠道微生物群在衰老过程中维持大脑健康和免疫力提供了证据。年轻小鼠供体的微生物群逆转了海马代谢产物和大脑免疫力中与衰老相关的差异,并在移植到老年宿主中时改善了与年龄相关的认知行为障碍。这些工作为使用益生菌、益生元制剂影响肠道微生物群以恢复免疫系统和衰老大脑提供了可能性。异时的粪便转移证实了微生物组成的年龄依赖性变化与宿主免疫系统功能下降之间的因果关系。事实上,老年小鼠派伊尔淋巴结中有缺陷的生发中心可以通过从年轻动物中FMT挽救,而不会影响外周淋巴结中的生发中心反应。最后,来自年轻供体小鼠的FMT改善了老年小鼠的卵巢功能和生育能力。这些有益作用与老年卵巢免疫微环境的改善有关,包括巨噬细胞和巨噬细胞衍生的多核巨细胞减少、促炎性IFNγ水平降低、抗炎细胞因子IL-4的丰度增加。
拓展:Blood:浙江大学钱鹏旭/黄河通过移植自年轻小鼠的粪便菌群通过抑制炎症使衰老的造血干细胞恢复活力
其他延长肠道微生物群的干预措施
益生菌植物乳杆菌GKM3在加速衰老的SAMP8小鼠模型中促进长寿并缓解与衰老相关的认知障碍。针对肠道微生物群组成的干预措施也恢复了小胶质细胞成熟和功能的衰老相关性下降,这会导致大脑可塑性变化并促进神经退化。肠道微生物群代谢产物如SCFA的再克隆实验或给药,防止了有益双歧杆菌的年龄相关性下降,增加了阿克曼氏菌的丰度,并恢复了中年小鼠的小胶质细胞功能。此外,热量限制饮食会导致肠道微生物组的结构变化,增加乳酸杆菌和其他影响健康衰老的物种的丰度。通过恢复老年小鼠和猕猴体内有益的SCFA产生细菌(嗜黏蛋白阿克曼菌)的丰度,肠道微生物引起的炎症和随后胰岛素抵抗的增加也可以逆转。同样,在超重/肥胖胰岛素抵抗志愿者中进行的一项随机、双盲、安慰剂对照的试验研究表明,口服巴氏杀菌法处理的嗜黏蛋白阿克曼菌改善了胰岛素敏感性,降低了胰岛素血症和血浆总胆固醇水平,这些结果强调了衰老和生物失调之间的因果关系,并表明恢复年轻微生物组的干预措施可以延长健康寿命和寿命。
总结
衰老的所有12个特征都彼此密切相关(图7)。例如,基因组不稳定性(包括由端粒缩短引起的不稳定性)与表观遗传改变(通过表观遗传修饰物如TET2的功能缺失突变)、蛋白稳态丧失(产生突变、错误折叠的蛋白)、巨自噬障碍(通过自噬的能力去除多余的中心体、核外染色质和胞质DNA)、营养感应失调(因为SIRT6是一种NAD+传感器,参与DNA修复,但也对营养缺乏作出反应)、线粒体功能障碍(mtDNA突变)、细胞衰老(DNA损伤触发衰老)、细胞间通讯的改变(通过阻碍通讯途径的激活)、慢性炎症(因为CHIP和DNA泄漏到胞浆中导致炎症)和生态失调(由于肠细胞中的突变有利于生态失调,而特定的微生物蛋白和代谢产物充当诱变剂)密切相关。类似的功能关系可以列出大多数而不是所有衰老特征,表明它们之间的强大相互联系。
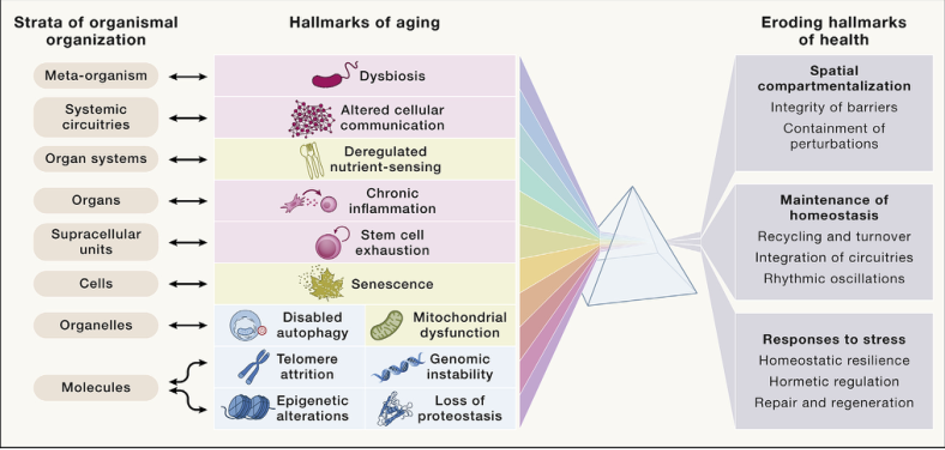
图7 本研究中提出的所有12个衰老特征在功能上相互关联。衰老的这些决定因素也与健康的八个特征相互关联,这八个特征包括空间划分的组织特征、随着时间的推移保持体内平衡以及对扰动的一系列反应。最后,衰老的特征与提出的八个组织层次相互关联,并创造了一个多维的交互空间,这可能有助于解释衰老过程的一些重要特征。
衰老特征之间这种密切关联在实验性延缓衰老干预措施的水平上也很明显,这些干预措施往往同时针对几个特征。因此,包括NAD+前体在内的SIRT激活剂减弱了基因组不稳定性(通过DNA修复)、表观遗传改变(通过组蛋白去乙酰化)、蛋白稳定性丧失(通过去除蛋白质聚集体)、巨自噬障碍(通过自噬增强)、营养感应失调(通过激活营养缺乏传感器)、线粒体功能障碍(通过线粒体吞噬依赖性质量控制的增加)。肠道细菌产生的代谢物亚精胺与DNA络合(抵消基因组不稳定性)、影响翻译(避免蛋白停滞)、刺激巨自噬、逆转淋巴细胞衰老、防止肌肉干细胞耗竭、维持昼夜节律、抑制炎症、刺激癌症免疫监测。二甲双胍具有多效性作用模式,包括诱导自噬、激活营养缺乏传感器AMPK、抑制线粒体呼吸、减缓脂肪细胞衰老、抑制炎症和促进肠道微生物群的有利变化。同样,通过口服补充嗜黏蛋白阿克曼菌维持生态平衡、刺激肠道自噬、减少代谢综合征、抑制炎症、增强抗癌免疫反应。事实上,降低胰岛素/IGF-1信号和破坏TORC1复合物等能够有效延缓衰老的干预措施的显著特征是具有针对不同组织不同衰老特征的多样性机制,从而维持整个机体健康寿命。
衰老的12个特征中的每一个都可以单独干预,从而延长健康寿命和寿命(表1),但它们之间并不平等(图1)。因此,正如作者最初所说的,主要特征反映了基因组、端粒、表观基因组、蛋白质组和细胞器等方面的损伤,这些损伤会随着时间积累,并直接推动衰老过程。对抗性特征则是指细胞对损伤的应答反应,在衰老过程中起着复杂的作用。例如,营养物质感应激活的营养信号和合成代谢反应在年轻时有利于生长发育,但在老年时则加速衰老。因此,在对抗性多效性的典型例子中,营养物质感应网络在个体成熟之前有益于器官形成,但在之后则变得有害。此外,线粒体功能障碍如果是可逆的和低剂量的,可以激发有益的逆反应(通过促分裂),而细胞衰老如果是有限的和局部的,可以帮助抑制肿瘤和促进伤口愈合。最后,当原发性和对抗性特征造成的损伤超出了细胞的补偿能力时,就会出现综合性特征,导致干细胞功能丧失、细胞间通讯紊乱,包括ECM破坏、慢性炎症和生态失调等。这些共同决定了衰老的速度。
最近,作者提出了健康的八个特征,包括组织的空间分区(屏障完好和局部干扰受限)、稳态长期保持(循环和更新、回路协调和节律同步)以及对干扰的适度反应(稳态恢复、兴奋调节、修复和再生)。显然,老化与健康的这八个特征的逐步衰退有关,这意味着越来越难以维持空间划分(伴随着内外屏障破坏和无法控制的时空干扰),保证长期稳态(伴随着循环和更新能力下降,不同系统之间通过“集成电路”的低效配合,以及昼夜节律或亚昼夜节律的失调),并通过完整修复和再生、稳态恢复和兴奋调节有效应对压力(图7)。这种衰退影响了所有八个器官组织层级,涉及不同类别的分子(如DNA、RNA、蛋白质和代谢物)、细胞器(如细胞核、线粒体和溶酶体)、细胞类型(如实质细胞、辅助/基质细胞和炎性/免疫细胞)、构成器官最小功能单元的超细胞结构、整个器官内的解剖边界、器官系统(如消化道、呼吸道和泌尿道)、系统回路(及其内分泌、神经、淋巴和血管连接)以及元有机体(包括微生物群)。结果表明,老化的12个特征与健康衰退的8个特征和组织的8个层级相互联系(图7),形成了一个多维的交互空间,可以解释老化过程中出现的一些现象。
年轻和年老小鼠血管系统连接在一起的异时异体共生实验可以最好地说明系统调节因素(如激素和循环细胞)在老化过程中的重要性。这种现象已经在单细胞转录组学水平上得到了广泛表征,揭示了年轻因子延缓老化因子加速年轻或老化细胞老化的时空图。这种类型的实验表明,老化依赖于细胞自主和非细胞自主机制的整合,在果蝇和小鼠中也有证据支持。因此,促老化和延缓老化机制可以在不同的细胞类型之间进行沟通,也许可以解释“正常”老化通常以接近同步的方式影响多个器官。而在“病理性”老化中,则会出现时间依赖性疾病,在特定位置过早地表现出心血管、肿瘤或神经退行性疾病。然而,“正常”和“病理性”老化之间的区别是有争议的,并且一些类激素综合征表现出不完全或节段性老化迹象,例如HGPS中缺乏中枢神经表型。
考虑到制定长寿策略方面取得了惊人进展(表1),制定干预人类老化的合理策略将非常重要。问题是,延长人类健康寿命战略应该在多大程度上针对避免加速老化环境因素(如污染、压力、体力活动不足和不健康的饮食,这些因素在贫困、不稳定和战时环境中往往不可避免)、采用促进健康生活方式因素(如饮食、锻炼、规律的睡眠模式和社交活动)、服用相对非特异性多效药物(例如NAD+前体、二甲双胍、精胺或MTORC1抑制剂等)或更具体医疗干预措施。这种特定治疗可能涉及药理学药物、基因或细胞疗法或生物工程方法等复杂逻辑和高成本手段。考虑到提供减缓、停止或逆转衰老的治疗策略的多重特征,评估联合疗法的最大效益和最小副作用将是有趣的。这种延长健康寿命和寿命的预防性治疗是否会受益于根据衰老时钟的基因组、表观基因组、代谢组或表型评估确定的个体患者个性化特征,这个问题仍然悬而未决。
衰老尚未成为药物开发或治疗的公认目标。因此,评估延缓衰老干预措施的第一个临床试验必须涉及预防或减轻与年龄相关的病理,而不是衰老本身。一个值得关注的问题是,尽管此类试验旨在针对高危人群(如CANTOS试验中的心肌梗死和实验室炎症症状患者,或未来二甲双胍相关试验中的虚弱或心血管事件患者),并为了测量继发性心血管事件或虚弱加重的表现,这类试验存在过晚干预或规划的风险。事实上,在这一点上,学术认知可能会随着第一次随机、双盲III期试验的结果而升高或降低。老龄化特征的新方向可能为制定旨在延长健康寿命的有效干预措施提供了一个改进的框架。
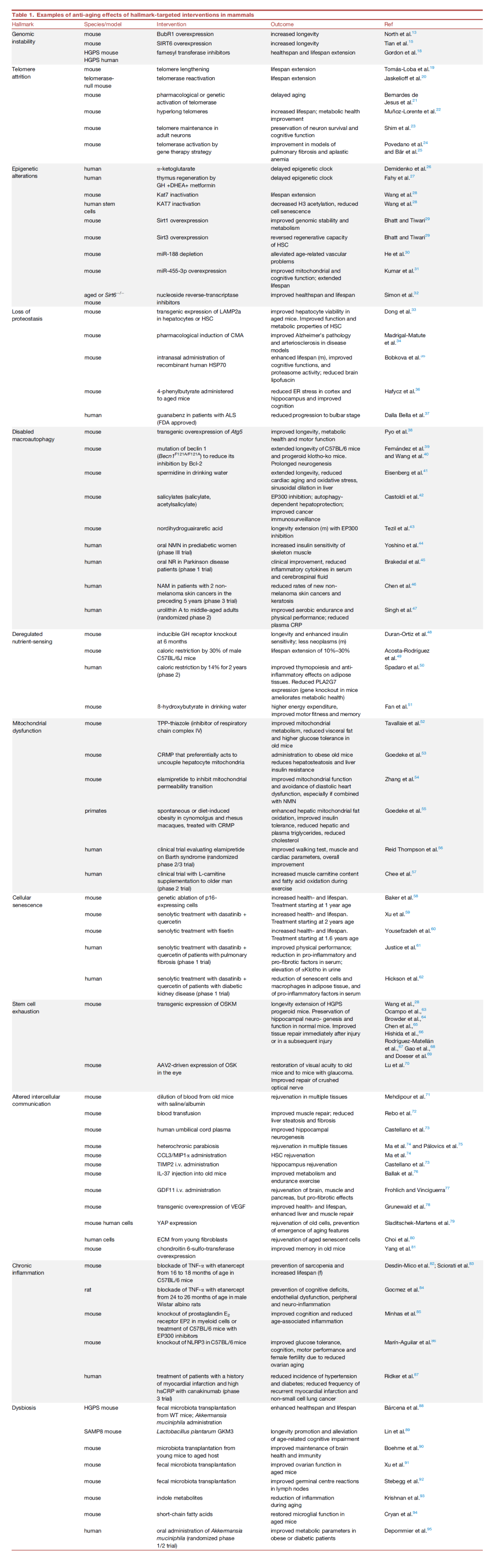
表1 哺乳动物中靶向衰老特征延缓衰老
 补充:
补充:
当然,除了这些分子因素,其实心理,运动同样重要,也值得深入研究。
拓展:
原始出处:
López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: An expanding universe.Cell . 2023 Jan 19;186(2):243-278. doi: 10.1016/j.cell.2022.11.001. Epub 2023 Jan 3.PMID: 36599349
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言









#衰老#还是许多常见疾病(如癌症、#糖尿病#、心脏病、神经退化等)的根源。幸运的是,科学家们在衰老领域取得了令人惊叹的进展,特别是发现了一些可以用来判断和评估衰老程度的特征和标志物。#二甲双胍##雷帕霉素##亚精胺##尿苷##自噬##基因组稳定性##表观遗传学##细胞衰老##humanin##AKK菌##Senolytics##生酮饮食##间歇性断食#,太多的关键词了,这些都与衰老有关,不过,从整体而言,心理平和(不#焦虑#),适度#运动#,合理膳食,再加上适当的#保健品#,及时进行疾病筛查和治疗,这可能是#抗衰老#的关键因素。
210