Nat Rev Neurosci 综述:突触功能的另一面
2022-11-29 brainnew神内神外 brainnew神内神外 发表于安徽省

突触前和突触后神经元末梢的分子组成是动态的,突触后反应的长期稳定对于突触发育和长期可塑性是必要的。由于突触的分子构成中与学习和记忆相关的可塑性变化,协调这些概念变得更加复杂。
突触前和突触后神经元末梢的分子组成是动态的,突触后反应的长期稳定对于突触发育和长期可塑性是必要的。由于突触的分子构成中与学习和记忆相关的可塑性变化,协调这些概念变得更加复杂。单颗粒追踪技术的进步意味着我们现在可以量化特定突触分子的数量和扩散特性,而统计热力学为分析这些分子波动提供了基础。
近日,Stephanie A. Maynard, Jonas Ranft & Antoine Triller在Nature Review Neuroscience 杂志上发表了最新的综述Quantifying postsynaptic receptor dynamics: insights into synaptic function。 突触是一种高度组织化的大分子结构,可确保动作电位从一个神经元传递到另一个神经元。动作电位到达突触前活性区导致神经递质从突触前囊泡释放,随后激活突触后受体靶标,这些靶标通过专门的支架蛋白聚集在突触后膜上(图1a)。
突触是一种高度组织化的大分子结构,可确保动作电位从一个神经元传递到另一个神经元。动作电位到达突触前活性区导致神经递质从突触前囊泡释放,随后激活突触后受体靶标,这些靶标通过专门的支架蛋白聚集在突触后膜上(图1a)。
以前,突触被视为一种静态结构,其突触后受体簇群的空间受限区域面向活动区 (the active zone)。然而,现在我们知道,尽管其结构高度有序,但突触的分子成分是动态的。
突触后受体组织
突触后末端是聚集在膜上并被支架蛋白限制的受体(图1a)。在兴奋性神经元中,这被称为“突触后密度”(PSD),在抑制性神经元中,它被称为“突触后域”或“突触后特化”。
受体分别通过胞吐和胞吞途径在突触外的位点传递到突触后膜并从突触后膜移除(图1a)。“扩散捕获”模型表明,在突触后膜的突触外部分扩散的受体通过与支架蛋白的瞬时相互作用或由于其他跨膜蛋白或细胞骨架元件造成的分子拥挤(图1b)。
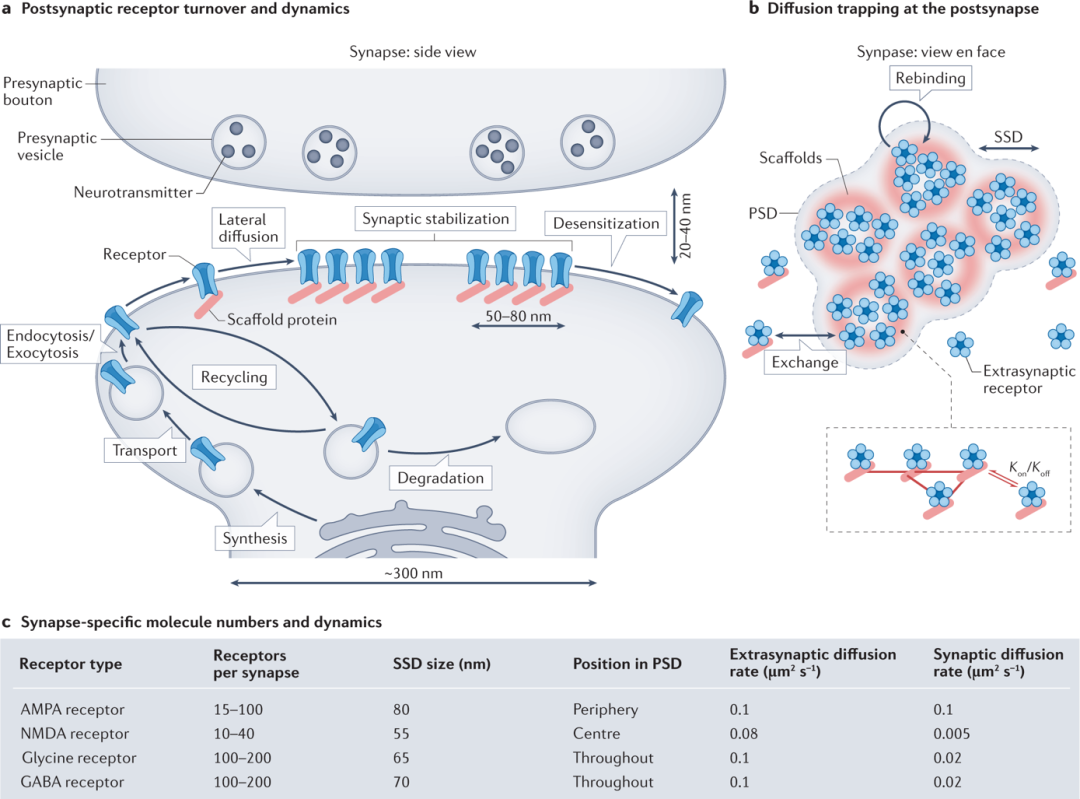 图1.突触处突触后受体的结构和动力学。
图1.突触处突触后受体的结构和动力学。
兴奋性和抑制性受体的组织
包含 NMDA 受体(NMDAR)的兴奋性突触形成每个突触的 10-40 个受体簇,这些受体也与 PSD95 结合(图1c)。树突棘 PSD 内的几个兴奋性突触后受体的空间组织已被证明与棘形态有关,将受体组织与突触功能联系起来。总的来说,这些发现表明突触内的分子组织(决定突触传递效率)受到高度调节。
兴奋性和抑制性受体的扩散系数
据报道,受体扩散系数在 0.005-0.1 μm2 s-1 的范围内,突触内和突触外扩散系数之间具有很强的变异性。抑制性受体表现出突触后膜横向扩散,其时间尺度与兴奋性受体的时间尺度相当(图1c)。
一个重要的问题是如何协调突触后受体的这些动态特性与维持持续突触身份、完整性和功能的需要,并允许与学习和记忆相关的可塑性变化。大型单分子荧光成像数据集已经能够通过统计热力学进行分析。这些分析可以提供无法直接测量的新观测值(图2),并允许我们对突触发生和突触可塑性等过程中受体扩散的局部波动进行定量分析,并将这些与稳态受体运动进行比较。
 图2. 超分辨单粒子轨迹分析。
图2. 超分辨单粒子轨迹分析。
分子运动的统计推断
从 SPT (single-particle tracking,单粒子跟踪)数据计算的最常见参数包括随时间变化的扩散系数(D)和均方位移 (MSD)(图2a)。现在已经开发了许多更复杂的方法来更准确地描述分子运动。推理方法评估不同运动模型以定量方式描述数据的可能性(图2d,e)。
翻译后修饰和泛素化
影响受体横向扩散
突触后反应中,活动的长期稳定,是造成长期可塑性的原因。与神经递质受体相关的蛋白质的翻译后修饰(post-translational modifications)正在成为突触侧向扩散和亲和捕获(affinity trapping:由于其与结合伙伴的亲和性,分子被困在特定的细胞位置)的主要调节因子,这些突触被认为是这些稳定的基础。
NMDAR 依赖性突触强度的长期抑制已被证明,通过使用基于 PAINT 的 SPT 成像,诱导突触处 PSD95 的去磷酸化,导致 AMPAR 表面扩散增加,突触 AMPAR 含量损失(图3a)并增加 CA1 神经元的短期可塑性。Radixin去磷酸化或消耗从突触外池中释放含有α5亚基的GABAAR,导致突触受体数量增加,和微型抑制性突触后电流的频率增加(图3a)以及短期记忆和逆转学习受损。
受体脱敏和构象影响流动性
短期可塑性依赖于神经递质受体的激活及其从脱敏中的恢复。脱敏受体与stargazin(脑特异性跨膜蛋白)的结合减少,从而降低了它们的突触稳定性。在突触前刺激后防止 AMPAR 扩散远离突触会降低兴奋性突触后电位幅度,从而降低短期可塑性(图3b)。因此,stargazin 在很长一段时间内调节突触部位可用的可激活 AMPAR 的数量(以及突触强度)。
受体迁移率随树突距离的不同而不同
这些受体在近端树突节段中的组织和扩散受到它们与 CaMKII 相互作用的调节(图3c),CaMKII 在近端树突棘中以更高的密度聚集。阻断神经元活动会增加近端节段中含有 GluN2B 的 NMDAR 数量,这表明近端节段中这些受体的低密度是由神经元活动维持的。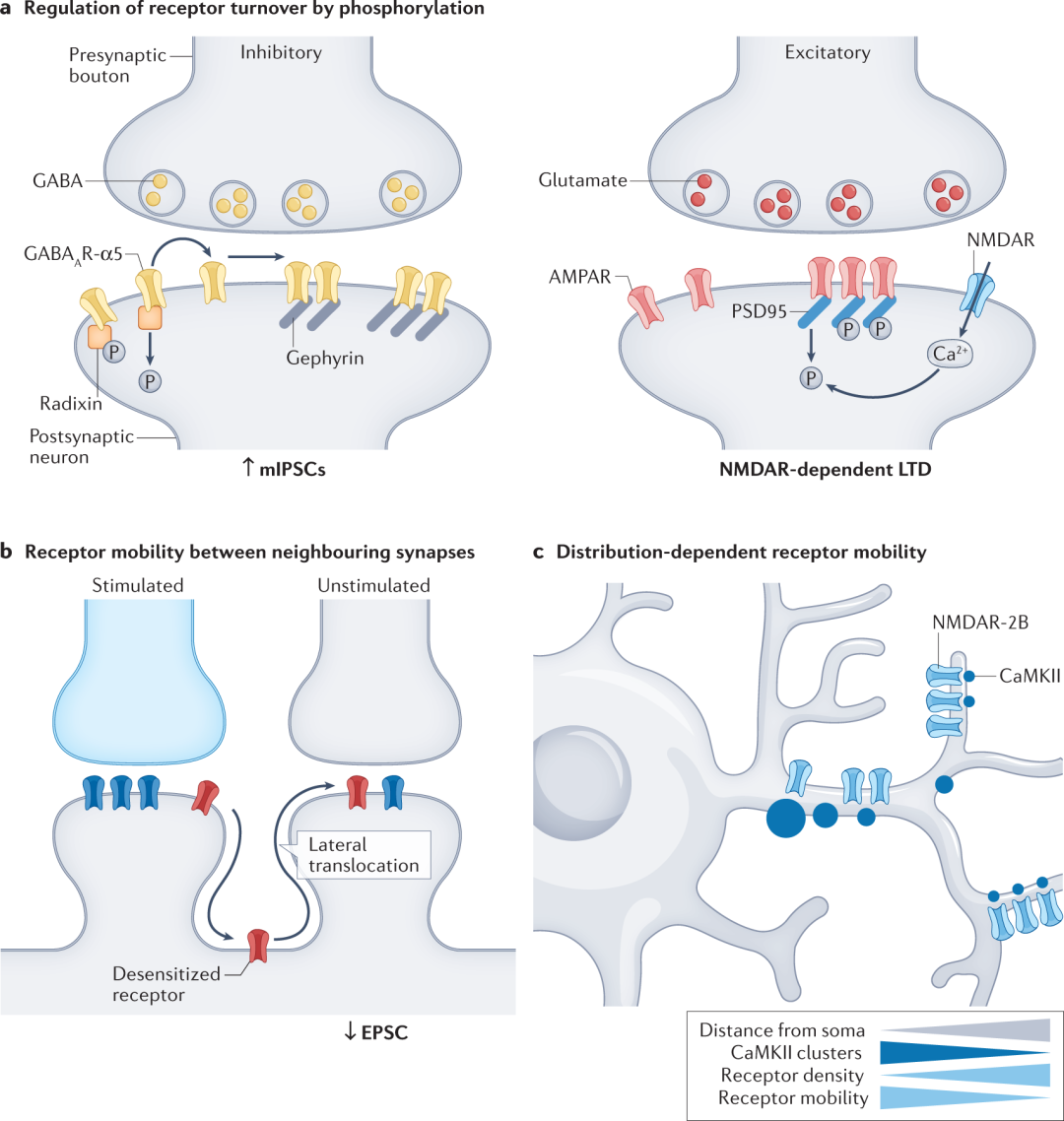 图3. 突触特异性时间动态。
图3. 突触特异性时间动态。
结 论
SPT 超分辨率显微镜、能够对轨迹类型进行分类和提取物理参数的跟踪算法,以及对这些高密度成像数据集进行建模,以推导出新的可观测数据的组合,这提供了时间分辨分子动力学的定量描述。
它们揭示了在分子水平上对突触传递的结构基础的重要见解,并使我们理解如何在更长的时间尺度上调和分子周转和突触稳定性。让我们在分子动力学上对突触反应、突触形成和可塑性的功能变异性的贡献有了新的认识。
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言




认真学习了
35